КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
X-1.3.4.2.2.2. Neolepidoptera Packard 1895
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Eumetabola
Eumetabola![]() Metabola
Metabola![]() Mecopteriformia
Mecopteriformia![]() Amphiesmenoptera
Amphiesmenoptera![]()
Lepidoptera![]() Glossolepidoptera
Glossolepidoptera![]() Neolepidoptera)
Neolepidoptera)
(рис. 10.16.2)
|
Название, неоднозначно соответствующие по циркумскрипции (см. выше, «Статус и классификация Palaeolepidoptera»): — Neolepidoptera Packard 1895. Иерархическое название: Papilio/fg5 [f:1781; g:L.1758(gen.203)] (sine Eriocrania; incl. Hepialus). |
|
Ранг. Инфраотряд или другой ранг между подотрядом и надсемейством.
Аутапоморфии Neolepidoptera. Помимо названной ниже особенности куколки, описаны аутапоморфии Neolepidoptera, связанные с тонкими деталями строения скелета, мускулатуры и внутренних органов имаго (Nielsen & Kristensen 1996).
(1) Склеенная куколка. Куколка не имеет подвижных мандибул («pupa adectica»); у подавляющего большинства Neolepidoptera куколка склеенного типа [«pupa conglutinata» – см. Metabola (1Bb)] – т.е. ее антенны, ротовые придатки, конечности и протоптероны плотно приклеены к туловищу и кажутся рельефом на поверхности, а поверхностная кутикула склеротизована (рис. 10.16.2E). Таким образом, для выхода из своего укрытия куколка не может пользоваться ногами и мандибулами, и может перемещаться только за счет движений брюшка. У разных Neolepidoptera укрытием служит либо кокон [сплетенный из шелка лабиальных желез – см. Mecopteriformia (3)], либо колыбелька (в почве, древесине и т.п.), либо укрытие отсутствует (например, у Rhopalocera).
У примитивных представителей Neolepidoptera куколка имеет относительно длинное подвижное брюшко, у которого подвижно сочлененными бывают I–VII уромеры; иногда I или I–II уромеры неподвижно слиты с грудью, иногда VII уромер неподвижно слит с VIII–X уромерами. У большинства представителей таксона Ditrysia брюшко куколки короткое, и подвижны только V и VI уромеры (рис. 10.16.2E); чешуекрылых с такими куколками иногда объединяют в таксон Obtectomera Minet 1986.
В отличие от Neolepidoptera, прочие Amphiesmenoptera (Palaeolepidoptera, Protolepidoptera и большинство Trichoptera) имеют куколку примитивного несклеенного типа с функционирующими мандибулами – «pupa dectica» [см. Amphiesmenoptera (10)].
Помимо Neolepidoptera, склеенные куколки возникли у двукрылых, у некоторых перепончатокрылых и у некоторых жуков; среди них, по крайней мере у перепончатокрылых и жуков склеенные куколки произошли от несклеенных куколок с нефункционирующими мандибулами («pupa adectica exarata») (см. Указатель признаков [5.24]); в отличие от них, у Neolepidoptera склеенная куколка произошла непосредственно от примитивной куколки «pupa dectica», без промежуточного этапа в виде «pupa adectica exarata» (Hinton 1946).
Признак Neolepidoptera неясного филогенетического значения
(2) Гусеница. Личинка представляет собой гусеницу (eruca): длинное червеобразное туловище опирается, помимо трех пар грудных ног [см. (2a)], на 4 пары брюшных ложноножек [см. (2b)] и на парный пигопод [см. (2c)]. Такое строение исходно для Neolepiodoptera; однако неясно, является ли оно исходным только для Neolepidoptera, или также для вышестоящих таксонов Glossolepidoptera и Lepidoptera [см. (2d)]. Строение тела гусеницы приспособлено прежде всего для ползанья по растениям, и большинство гусениц является фитофагами [см. Lepidoptera (6)].
(2 a) Грудные ноги обычно сильно укорочены, но могут иметь все членики – тазик, вертлуг (обычно сильно укороченный и б.м. слитый с бедром), бедро, голень, лапку [одночлениковую – см. Metabola (1Aa)] и коготок [один – см. Mecopteriformia (4)] (рис. 10.16.2C). Ноги почти всегда короткие, очень редко вторично удлинены; иногда ноги вестигиальные или совсем утрачены (что часто бывает связано со специализацией к обитанию внутри тканей растений и сопровождается одновременной утратой ложноножек). В отличие от других Metabola, у изученных представителей Neolepidoptera личиночная нога трансформируется в куколочную без фазы дегенерации (рис. 10.16.2D) [см. Metabola (1Ccδ-12)].
(2 b) Ложноножки расположены по паре на вентральной стороне III–VI сегментов брюшка; на I–II и VII–IX сегментах брюшка ложноножки отсутствуют (рис. 10.16.2A). Пара рядов ложноножек расположена как продолжение пары рядов грудных ног [см. Metabola (1Abβ)]. В типичном случае каждая ложноножка представляет собой цилиндрический вырост вентральной стороны тела; подошва ложноножки (planta) окаймлена кольцевидным рядом склеротизованных крючков, изогнутых остриями кнаружи. От боковой стенки соответствующего сегмента к подошве ложноножки идут мышцы, гомологичные дорсовентральным мышцам обычного брюшного сегмента (рис. 10.16.2A); при сокращении этих мышц подошва ложноножки складывается и втягивается внутрь, ложноножка укорачивается, а крючки сближаются и поворачиваются остриями дистально; обратное движение – удлинение ложноножки, распрямление подошвы и поворот крючков остриями наружу происходит под действием давления гемолимфы при сокращении туловищных мышц.
Ложноножки такого расположения и строения характерны для большинства, но не для всех Neolepidoptera (Hinton 1955). У многих гусениц ряд крючков не образует цельного кольца и развит только на той стороне подошвы, которая обращена медиально. У некоторых гусениц число ложноножек уменьшено: некоторые из передних ложноножек бывают утрачены, так что ложноножки имеются на IV–VI или на V–VI уромерах (у некоторых совок – Noctua/fg и др.), либо лишь на VI уромере (у пядениц – Geometra/fg); у некоторых гусениц, наоборот, утрачены ложноножки VI уромера. У некоторых гусениц, специализированных к обитанию внутри растений, а также у открытоживущих слизневидок (Limacodes/f=Apoda/g) все ложноножки, а иногда и грудные ноги, полностью утрачены. Увеличение числа ложноножек произошло у гусениц Megalopyge/fg, у которых на II и VII уромерах имеется по паре дополнительных ложноножек без крючков.
(2 c) Пигопод [см. Metabola (1Abα)] представляет собой пару направленных вентрально анальных ложноножек на конце последнего, X уромера (рис. 10.16.2A). Эти анальные ложноножки напоминают ложноножки III–VI уромеров, но отличаются от них тем, что кольцо крючков всегда не замкнуто сзади, а число мышц, идущих к подошве, больше. У некоторых гусениц анальные ложноножки утрачены и имеются лишь ложноножки на III–VI сегментах брюшка.
(2 d) Статус признака. Ложноножки личинок независимо возникали в разных таксонах насекомых с полным превращением (см. Metabola (1Abβ) и Указатель признаков [3.44]); такие же по строению ложноножки, как у Neolepidoptera, имеются у некоторых двукрылых, но там они имеют иное число и расположение [ср.: Diptera (3)].
Нет сомнений в том, что характерное строение гусениц с крючконосными ложноножками на III–VI и X уромерах возникло однократно и является аутапоморфией. Неясно только, аутапоморфией какого именно таксона оно является – Neolepidoptera, Glossolepidoptera или Lepidoptera в целом. У всех чешуекрылых кроме Neolepidoptera личинки специализированы к скрытому образу жизни, и их органы передвижения подверглись редукции. Среди них только личинки Micropteryx/fg (incl. Sabatinca) активно передвигаются в подстилке, но их ноги сильно редуцированы, а на брюшке имеются ложноножки принципиально иного строения и расположения, чем ложноножки гусениц Neolepidoptera. Все остальные известные личинки Protolepidoptera и Palaeolepidoptera обитают в тканях растений и не имеют ложноножек (ср.: Protolepidoptera: «Статус и классификации» и Palaeolepidoptera: «Статус и классификации»); неясно, является ли отсутствие ложноножек у них первичным или это результат редукции, поскольку те личинки Neolepidoptera, которые специализировались к обитанию внутри растений, тоже не имеют ложноножек.
Размеры. От мелких (от 3 мм) до крупных (до 30 см в размахе крыльев).
Геологический возраст. Юра – ныне (Rasnitsyn & Quicke 2002).
Состав и распространение. Более 150 000 видов (Zhang 2011); всесветно.
Классификации Neolepidoptera
I. Деление Neolepidoptera на Exoporia и Frenatae. Согласно наиболее широко принятой точке зрения, филогения и классификация Neolepidoptera представляются следующим образом:
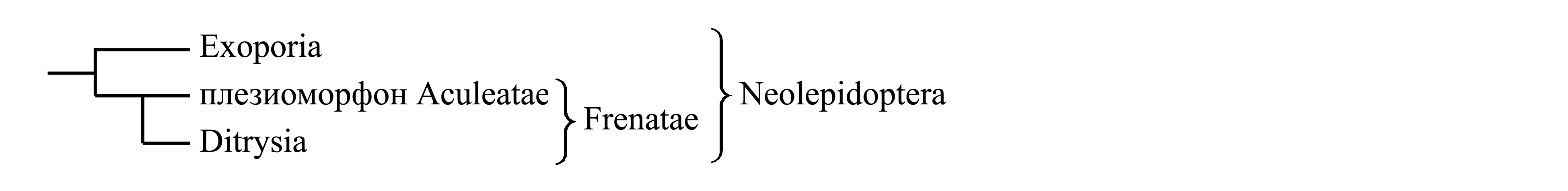
1. Exoporia (Dugdale 1974) Common 1975 {«exoporian Ditrysia» sensu Dugdale 1974; nom.hier.: Hepialus/fg1 [f:1809; g:1775] (incl. Mnesarchaea)}. У самки [не имеющей первичного яйцеклада – см. Amphiesmenoptera (9)] вместо одного генитального отверстия имеется два непарных отверстия – яйцекладное и копулятивное; оба отверстия находятся близко друг к другу позади VIII стернита брюшка; яйцекладное отверстие ведет в яйцевод; копулятивное отверстие находится вентральнее и ведет в копулятивную сумку; снаружи копулятивное и яйцекладное отверстия могут быть соединены желобом, по которому сперма из копулятивного отверстия попадает на яйца. Таким образом, по строению женских гениталий Exoporia напоминают Ditrysia (= Endoporia) (см. II). Exoporia делятся на Mnesarchaea/fg и Macrojugatae.
1.1. Mnesarchaea/fg [f:1912; g:1885]. Мелкие (8–12 мм) молевидные бимоторные бабочки с функционирующим хоботком. Несколько видов в Новой Зеландии.
1.2. Macrojugatae Comstock 1893 {synn.circ.: Jugata Tillyard 1918b (non Jugatae Comstock 1892)} = Aplostomatoptera Kiriakoff 1948 = Gymnocera Friese 1970; nom.hier: Hepialus/fg2 [f:1809; g:1775] (incl. Prototheore)}. Мелкие и крупные (10–100 мм) бимоторные бабочки с редуцированным ротовым аппаратом. Более 500 видов.
2. Frenatae Comstock 1892 {synn.circ.: Anisoneuria Karsch 1898 = Heteroneura Tillyard 1917d (non Heteroneura Brauer 1885); nom.hier.: Papilio/fg6 (sine Hepialus; incl. Nepticula, Adela)}. Включает подавляющее большинство видов Lepidoptera. Характеризуется переднемоторностью с уникальным строением крыловой сцепки. У самца френулюм [исходно представляющий собой группу щетинок – см. Metabola (4)] цельный, образован слитыми щетинками; начинаясь от основания переднего края заднего крыла, френулюм дугообразно тянется вперед и дистально; на вентральной стороне переднего крыла имеется особого строения зацепка, или ретинакулум (retinaculum) – отросток в виде крюка, направленный вентрально-назад и загнутый дорсально-вперед (по направлению к поверхности крыла); этот отросток находится либо на мембране между C и Sc, либо на жилке RA. В отличие от самца, у самки френулюм обычно состоит из нескольких неслившихся щетинок, а роль зацепки выполняет густой пучок чешуек, который расположен на вентральной стороне кубитальной жилки (Cu) переднего крыла и направлен вперед. У некоторых Frenatae самец имеет обе зацепки одновременно – и вырост на RA, и пучок чешуек на Cu.
У некоторых Frenatae френулюм утрачен. В этих случаях крылья либо не сцепляются, либо имеют иные сцепочные механизмы. У дневных бабочек (Rhopalocera), сатурний (Saturnia/fg) и некоторых других представителей таксона Frenatae сцепка крыльев является амплексиформной, т.е. обеспечивается лишь широким налеганием переднего крыла на заднее [см. Pterygota (4F)]; для этой заведомо полифилетической группы бабочек было предложено название Amplexiformia Tillyard 1918b. В некоторых случаях сцепка обеспечивается модифицированными чешуйками, расположенными около заднего края переднего крыла и около переднего края заднего крыла. Некоторые представители Frenatae, утратив френулюм, вторично перешли к бимоторности.
Помимо модификации френулума, таксон Frenatae характеризуется изменением заднего крыла; жилка RS на заднем крыле всегда неветвящаяся (тогда как у прочих чешуекрылых она имеет 3 или 4 ветви, как и на переднем крыле – рис. 10.16.1). В связи с переднемоторностью у многих Frenatae передние и задние крылья приобрели и другие различия. Заднее крыло может быть значительно уменьшено или, наоборот, расширено, с большой инверталярной областью, которая в покое переворачивается и складывается веерообразно.
Frenatae делят либо на Aculeatae и Ditrysia, либо на Nannolepidoptera и Eulepidoptera.
2.1. Плезиоморфон Aculeatae Spuler 1910 {orig.: «Tineides aculeatae» Spuler 1910; synn.circ.: Aculeata Fracker 1915 = Monostomiogyna Le Marchand 1945; nom.typ.: Adela/fg [f:1850; g:1802] (incl. Nepticula)}. Объединяет всех Frenatae, не относящихся к Ditrysia.
По другой классификации голофилетический таксон Nannolepidoptera Kiriakoff 1848 {nom.typ.: Nepticula/fg [f:1854; g:1843]} относится не к плезиоморфону Aculeatae, а противопоставляется таксону под названием Eulepidoptera Kiriakoff 1848 (non Eulepidoptera Crampton 1916a), который, в свою очередь, делится на Adela/fg и Ditrysia.
2.2. Ditrysia Börner 1925 {syn.circ.: Distomyogyna Le Marchand 1945 = Endoporia auct. («endoporian Ditrysia» sensu Dugdale 1974); nom.hier: Papilio/fg7 (sine Nepticula, Adela)} включает подавляющее большинство видов чешуекрылых. Аутапоморфией Ditrysia является особое строение гениталий самки. У самки [не имеющей первичного яйцеклада – см. Amphiesmenoptera (9)] вместо одного генитального отверстия имеется два непарных отверстия – яйцекладное и копулятивное; яйцекладное отверстие смещено назад, иногда совмещено с анальным (как и у многих других бабочек) и ведет в яйцевод и семяприемник; копулятивное отверстие вторично смещено вперед, на сочленение VII и VIII стернитов брюшка, и ведет в копулятивную сумку, куда при копуляции попадает сперма; внутри тела проходит семенной проток, соединяющий копулятивную сумку с непарным отделом яйцевода. По этому протоку сперма попадает из копулятивной сумки сначала в яйцевод, а оттуда – в семяприемник; оплодотворение яиц, как и у других бабочек, происходит благодаря тому, что при прохождении яиц по яйцеводу сперма выдавливается из семяприемника обратно в яйцевод. В некоторых старых классификациях таксон Ditrysia противопоставлялся всем прочим чешуекрылым (см. Lepidoptera: «Классификации» III). По строению женских гениталий Ditrysia сходны с Exoporia, что может свидетельствовать о родстве Ditrysia и Exoporia (см. II).
II. Единое происхождение Exoporia и Ditrysia. Высказано предположение, что сходство в строении гениталий самки у Exoporia и Ditrysia обусловлено их единым происхождением (Dugdale 1974). В таком случае филогения Neolepidoptera должна выглядеть следующим образом:

И у Exoporia, и у Ditrysia копулятивная сумка расположена вентральнее яйцевода и открывается наружу копулятивным отверстием, отдельным от яйцекладного отверстия (тогда как у всех прочих Amphiesmenoptera копулятивная сумка располагается дорсальнее яйцевода и вместе с яйцеводом открывается наружу единым половым отверстием). У Exoporia сперма попадает из копулятивного отверстия в яйцекладное по открытому желобу. У Ditrysia этот желоб замкнулся и превратился во внутренний канал (см. I). Это предположение о едином происхождении двух половых отверстий противоречит представлению о голофилии Frenatae (см. I).
* * *