КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
IX-1.2.2. Condylognatha Börner 1904
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Eumetabola
Eumetabola![]() Parametabola
Parametabola![]() Acercaria
Acercaria![]() Condylognatha)
Condylognatha)
(рис. 9.4.1C, 9.4.2–9.12.4)
|
Названия, однозначно соответствующие по циркумскрипции: — Hemiptera Linnaeus 1758 (non auct.); — Proboscidea Scopoli 1763 (non Proboscidea Latreille 1809, nec Proboscidea Illiger 1811); — Reamurii Scopoli 1777; — Hemimeroptera Clairville 1798; — Elytroptera-Siphonostoma Latreille 1807 (non Gymnoptera-Siphonostoma); — Hemipteria Rafinesque 1815; — Condylognatha Börner 1904a; — Condylognathida Boudreaux 1979b. По циркумскрипции также соответствует таксонам под типифицированными названиями, образованными от родового названия Cimex, в частности Cimicoides (sensu Laicharting 1781). Иерархическое название: Cimex/f1=Cicada/g3 [f:1781(ord.3); g:L.1758(gen.195)] (sine Pediculus; incl. Thrips). |
|
Ранг. В XX веке – обычно надотряд и выше; в XVIII и начале XIX века – отряд.
Название. Наиболее старым названием, соответствующим по своей исходной циркумскрипции этому таксону, является широко известное название Hemiptera, которое в соответствии с исходной датой приоритета (см. I.6.3.1.2 и I.6.5.2) имеет авторство «Linnaeus 1758». Однако в современной литературе название «Hemiptera» большей частью употребляется в других значениях – дибо для Arthroidignatha (см. ниже), либо для Heteroptera, так что использование его для данного таксона может вызвать путаницу. Забытое название Proboscidea Scopoli 1763 имеет широко известный младший омоним – Proboscidea Illiger 1811 (слоны); кроме того, такое же название Proboscidea Latreille 1809 присваивалось таксону в составе Diptera; такое же название присваивалось и некоторым другим таксонам животных. В настоящее время наиболее принятым названием этого таксона является Condylognatha Börner 1904. Его следует считать однозначно соответствующим по циркумскрипции данному таксону, несмотря на то, что исходно к Condylognatha был отнесен, помимо прочего, таксон Conorrhyncha (см. выше, о названии Acercaria).
Аутапоморфии Condylognatha
(1) Колюще-сосущий ротовой аппарат. Ротовой аппарат имеет уникальное строение: трубчатый внутренний хоботок (proboscis interior) образован сцепленными стилетами максиллярных лациний [см. (1b)], способен выдвигаться, проникать в субстрат и проводить жидкую пищу; мандибулы преобразованы в колющие стилеты [см. (1a)]; гипофаринкс образует слюнной шприц [см. (1c)]; верхняя губа, стипесы максилл и нижняя губа плотно сомкнуты и образуют ротовой конус [см. (1d)], из вершины которого выдвигаются максиллярные и мандибулярные стилеты.
Вероятно, исходно такой ротовой аппарат адаптирован для высасывания мицелия грибов (как у многих современных Thysanoptera) и/или высасывания клеточного содержимого и соков из зеленых частей сосудистых растений (как у многих современных Thysanoptera, у всех Plantisuga, Auchenorrhyncha, Coleorrhyncha и многих Heteroptera). Этот же ротовой аппарат может служить и для хищничества (у некоторых Thysanoptera и Heteroptera), питания пыльцой (у многих Thysanoptera), кровью позвоночных животных (у некоторых Heteroptera) и другой пищей.
Ниже (1a–d) ротовые части условно описаны так, как они расположены при гипогнатном положении головы (когда ротовые придатки направлены вершинами вентрально, клипеус и верхняя губа находятся спереди, а нижняя губа сзади). Такое положение действительно встречается, но в ряде таксонов (Neothysanoptera, Saltipedes, Auchenorrhyncha, Coleorrhyncha и др.) голова имеет опистогнатное положение (т.е. ротовые придатки направлены вершинами назад, а клипеус и верхняя губа находятся вентрально), а в других случаях (у многих Heteroptera) голова прогнатная (т.е. ротовые придатки направлены вершинами вперед, клипеус и верхняя губа находятся дорсально, а нижняя губа вентрально).
(1 a) Модификация мандибул. Мандибулы специализированы как колющие и скрыты внутри головы. Каждая мандибула сильно вытянута начиная от самого своего основания, а ее дистальная часть представляет собой тонкий заостренный мандибулярный стилет. Сочленение мандибулы с головной капсулой сохранят наружное положение, но вся мандибула вдавлена в голову так, что погружена в глубокий парный карман – мандибулярный футляр; исходно мандибулярный футляр спереди ограничен клипеусом, а сзади – стипесом максиллы; клипеус и стипес сомкнуты своими краями [см. (1d)], и щель между этими краями ведет в полость мандибулярного футляра (рис. 9.5.2) [у членистохоботных между клипеусом и мандибулярным футляром добавляется мандибулярная пластинка – см. ниже, Arthroidignatha (1a)].
Мандибулярные стилеты тянутся внутри своих футляров параллельно погруженным в голову максиллярным стилетам [см. (1b)]. На прозрачном препарате мандибулярные стилеты кажутся сходными с максиллярными стилетами; однако принципиальное отличие мандибулярного стилета состоит в том, что он погрузился не основанием, а стороной, так что его футляр открыт наружу щелью, тянущейся параллельно стилету, тогда как максиллярная лациния погружена в голову основанием, и ее футляр имеет вид трубки [см. Acercaria (1)].
У трипсов единственная функционирующая мандибула [левая – см. ниже, Thysanoptera (2)] не имеет мышц-протракторов (от ее основания отходят лишь мышцы, идущие дорсально к головной капсуле и способные служить ретракторами); для покалывания субстрата мандибулой трипсы либо наклоняют голову, либо выдвигают мандибулу опосредованным действием мышцы, идущей от стенки мандибулярного футляра к фронтоклипеусу [см. ниже, Thysanoptera (2)]. У членистохоботных мандибула претерпела дальнейшую модификацию, вошла в состав внутреннего хоботка, расчленена на стилет и рычаг и имеет протрактор, идущий к особой мандибулярной пластинке головы [см. ниже, Arthroidignatha (1a)].
Название «Condylognatha» исходно связано с недоразумением: его автор принимал лацинии и рычаги максилл трипсов [см. (1b)] за мандибулы и поэтому ошибочно считал, что у всех Condylognatha мандибула имеет отчлененный рычаг.
Поскольку полость мандибулы сильно сужена, перед линькой новая мандибула закладывается проксимальнее старой; у Arthroidignatha пара новых мандибул закручивается в полости головы такими же плоскими спиралями, как и пара максиллярных лациний [см. Acercaria (1с)] (Weber 1929).
(1 b) Модификация максилл. Лацинии максилл [втягивающиеся в голову – см. Acercaria (1)] преобразованы в колюще-сосущие максиллярные стилеты (рис. 9.4.1С, 9.5.2). Каждая лациния имеет желоб на медиальной стороне, так что складываясь и сцепляясь вместе, левая и правая лацинии образуют внутренний хоботок – единую трубку с каналом, по которому всасывается жидкая пища, а в противоположном направлении идет слюна [см. (1c)]; у трипсов имеется единственный канал, по которому пища и слюна могут идти только поочередно; у членистохоботных канал разделен на два – для пищи и для слюны [см. ниже, Arthroidignatha (1b) и рис. 9.7.2C]. Два максиллярных стилета способны сцепляться между собой благодаря тому, что на сторонах, обращенных друг к другу, они имеют комплементарные продольные гребни и желобки. Стилеты сцепляются друг с другом по мере выдвижения через узкое отверстие на вершине ротового конуса [см. (1d)] и расцепляются по мере втягивания в голову, где они разделены гипофаринксом [см. (1c)]. Таким образом, находящиеся внутри головы основания стилетов всегда расставлены, а дистальные части стилетов сомкнуты на том протяжении, на которое они выдвинуты за пределы ротового конуса.
Проксимальный конец лацинии подвижно сочленен с максиллярным рычагом; этот рычаг представляет собой склеротизованную внутреннюю аподему, начинающуюся от дна лациниального кармана и идущую сквозь миксоцель стипеса к его внешней стенке. Конец максиллярного рычага подвижно прикреплен изнутри к стенке стипеса (у Thysanoptera-Terebrantia) или соответствующей ему максиллярной пластинки [см. ниже, Arthroidignatha (1b)]. У имаго Thysanoptera-Tubulifera место прикрепления рычага сместилось со стипеса на головную капсулу. У Arthroidignatha-Hemelytrata максиллярная пластинка имеет аподему, слитую внутри головы с крыловидной аподемой гипофаринкса [см. ниже, Arhtroidignatha (1c)], и максиллярный рычаг крепится к этой эндоскелетной конструкции.
Максиллярная мала [см. Acercaria (1)] утрачена, так что максилла утратила способность загребать пищу в рот (в отличие от сеноедов и пухоедов, которые с помощью максиллярных мал перемещают в рот частицы твердой пищи, разгрызенной мандибулами).
(1 c) Модификация гипофаринкса. Гипофаринкс преобразован для проведения слюны в канал, образованный лациниями максилл [см. (1b)]: латеральные края гипофаринкса завернуты назад (т.е. по направлению к нижней губе) так, что между ними имеется продольный желоб, тянущийся до вершины гипофаринкса; в основание этого желоба открывается непарный проток слюнных желез [который у большинства других насекомых открывается между основанием гипофаринкса и нижней губы – см. Hexapoda (3)]; мышцы, расположенные внутри гипофаринкса, крепятся к стенке желоба, способствуя его расширению. У Thysanoptera желоб открыт назад, и слюнной канал образован им и нижней губой, прижатой к гипофаринксу сзади; у Arthroidignatha слюнной канал сместился внутрь гипофаринкса и имеет слюнной шприц особого строения [см. ниже, Arthroidignatha (1c)].
(1 d) Ротовой конус. Четыре придатка – верхняя губа, пара максиллярных стипесов [утративших кардо – см. Acercaria (1)] и нижняя губа – неподвижно торчат и сомкнуты своими краями так, что образуют единый выступ – ротовой конус (рис. 9.5.2). С каждой стороны ротового конуса латеральный край верхней губы плотно прижат к переднему краю максиллярного стипеса, тем самым закрывая сбоку мандибулярный карман, в котором скрыт мандибулярный стилет [см. (1a)], и от которого вглубь головы вдается максиллярный футляр с выдвигающейся из него максиллярной лацинией [см. (1b)]. Благодаря такому смыканию ротовых частей выдвигающиеся стилеты – лацинии максилл и мандибулы – выдвигаются только через узкий проход на вершине ротового конуса, так что при выдвигании они смыкаются и прижимаются друг к другу, образуя внутренний хоботок [см. (1a) и (1b)]. У Thysanoptera и некоторых Arthroidignatha вершина верхней губы свернута в трубку, образующую продолжение ротового конуса, через которое выдвигаются стилеты (рис. 9.5.2). Кроме того, у Arthroidignatha нижняя губа превращена в членистый внешний хоботок и образует продолжение ротового конуса, направляющее выдвигающиеся стилеты [см. ниже, Arthroidignatha (1) и рис. 9.6.1].
Признаки Condylognatha неясного филогенетического значения
(2) Парамеры. Гениталии самца [лишенные стерностилей – см. Eumetabola (4)] снабжены парой подвижных одночлениковых придатков, служащих для удержания брюшка самки. Эти придатки называют либо парамерами (предполагая их аналогию или гомологию с истинными парамерами жуков и других Metabola) (Snodgrass 1957), либо гоностилями (предполагая их аналогию или гомологию с истинными гоностилями Ephemeroptera). Далее мы будем называть эти придатки парамерами. Истинные парамеры насекомых являются частями фаллического комплекса и причленены между IX и X стернитами брюшка [см. Amyocerata (6)], тогда как истинные гоностили (гомологи стерностилей) причленены к заднему краю IX стернита брюшка или к стилигеру [см. Hexapoda (7)]. Отличить парамеры от гоностилей бывает трудно: из-за модификаций IX уромера места их причленения могут быть неразличимы; и к гоностилям, и к фаллическому комплексу могут подходить мышцы от IX стернита брюшка [см. и Amyocerata (6)]. Та же проблема гомологии пары хватательных генитальных придатков самцов касается и насекомых с полным превращением [ср.: Metabola (6)].
Парамеры Condylognatha причленены либо к основанию пениса, либо к заднему краю IX стернита брюшка, либо занимают промежуточное положение; мышцы к ним идут либо только от IX стернита брюшка (в этом случае парамеры можно принять за гоностили), либо от IX стернита и от основания пениса (в этом случае фаллическое происхождение парамеров очевидно). В отличие от парамеров Panpsocoptera, парамеры Condylognatha имеют мышцы, идущие от IX стернита брюшка [ср.: Panpsocoptera (2)]. В отличие от парамеров насекомых с полным превращением, парамеры Condylognatha всегда одночлениковые, без базипарамеров; мышцы крепятся к их основаниям и не заходят внутрь парамеров [ср.: Metabola (6)].
У Thysanoptera-Terebrantia, Auchenorrhyncha и Coleorrhyncha парамеры причленены к основанию пениса и, помимо мышц, идущих к IX стерниту брюшка, имеют мышцы, идущие к основанию пениса (рис. 9.4.2A–B); так что происхождение этих парамеров из обособившихся частей фаллического комплекса не вызывает сомнений. У листоблошек парамеры причленены к IX стерниту (так что напоминают гоностили), но соединены с основанием пениса посредством особого Y-образного склерита, имеющего собственные мышцы (рис. 9.4.2D) [см. ниже, Saltipedes (7)]; вероятно, Y-образный склерит, как и парамеры, является обособленной частью фаллического комплекса. У клопов парамеры не связаны с пенисом ни сочленениями, ни мышцами (рис. 9.4.2C) [см. ниже, Heteroptera (10)], так что по строению принципиально не отличаются от гоностилей. Однако парамеры Heteroptera имеют сходство с парамерами Auchenorrhyncha и Coleorrhyncha [см. ниже, Hemelytrata (7)], что говорит об их едином происхождении; синапоморфии Heteroptera и Coleorrhyncha [см. ниже, Heteropteroidea (1)–(3)] свидетельствуют о том, что единый план строения гениталий, свойственный Auchenorrhyncha и Coleorrhyncha, является исходным для Heteroptera.
В некоторых таксонах Condylognatha парамеры в той или иной мере редуцированы или полностью утрачены.
Среди трипсов парамеры сохраняются у Terebrania, но полностью утрачены у Tubulifera (см. ниже, Neothysanoptera: «Классификация»). Парамеры развиты у большинства цикадовых и клопов, но утрачены у певчих цикад и у некоторых клопов [см. ниже, Hemelytrata (7) и Heteroptera (10)]. Среди Plantisuga парамеры развиты у всех листоблошек и белокрылок [см. ниже, Saltipedes (7) и Scytinelytra (9)]. У тлей (Gynaptera) парамеры уменьшены, но могут сохранять подходящие к ним мышцы (Wieczorek et al. 2011: Fig. 1e); у многих тлей парамеры неподвижно слиты или утрачены. У кокцид парамеры полностью утрачены [см. ниже, Gallinsecta (7)].
(3) Тенденция к изменению метаморфоза. Число возрастов нелетающей стадии [см. Pterygota (4C)] сокращено до пяти или четырех: 5 возрастов у Thysanoptera-Tubulifera, большинства Hemelytrata и Saltipedes; 4 возраста у большинства Thysanoptera-Terebrantia, Scytinelytra, большинства Gynaptera и самцов Gallinsecta, а также у отдельных видов Hemelytrata-Heteroptera и Saltipedes; у некоторых Gynaptera число возрастов сокращено до 3 [см. Pterygota (4Ca) и Указатель признаков [5.16]: табл. G].
В нескольких неродственных друг другу таксонах в составе Condylognatha метаморфоз усложнен и имеет те или иные черты сходства с метаморфозом Metabola (т.е. с полным превращением): иногда изменяется расчленение ног [помимо увеличения числа тарсомеров с 2 до 3 – см. Parametabola (5)]; у трипсов (Thysanoptera), тлей (Gynaptera), кокцид (Gallinsecta) и белокрылок (Scytinelytra) происходит замена личиночных глаз на имагинальные; у кокцид, белокрылок и некоторых других личинка резко отличается от имаго; у трипсов и кокцид имеются неактивные стадии [см. ниже, Thysanoptera (1), Aphidococca (1), Gynaptera (2), Gallinsecta (1), Saltipedes (1), Scytinelytra (1)]. В двух неродственных таксонах – Thysanoptera и Gallinsecta – возник сходный тип метаморфоза: питающимися являются только личинки I и II возрастов, а следующие два или три возраста неактивные. Совокупность таких модифицированных типов метаморфоза иногда называют неометаморфозом, или неометаболией (neometamorphosis, neometabolia) (Weber 1933). Однако при этом неизвестны какие-либо модификации метаморфоза, которые были бы общими или исходными для всех Condylognatha.
Геологический возраст. Пермь – ныне (Rasnitsyn & Quicke 2002).
Систематическое положение Condylognatha. По хорошо сохранившимся инклюзам в бирманском янтаре, датируемым началом позднего мела, и по остаткам более раннего возраста, начиная с пермского, описаны взрослые самцы и самки, относимые к таксону Tetrastigmoptera Kluge 2019 {nom.typ.: Archipsylla/fg [fg:1906] или Permopsocus/fg [fg:1926]}. Это мелкие насекомые (2,5–5 мм); ротовые придатки (верхняя губа, мандибулы, максиллы и ментум нижней губы) удлинены, но при этом ротовой аппарат сохраняет исходную грызущую специализацию; максиллярные щупики 4-члениковые, лабиальные щупики 3-члениковые; обе пары крыльев сходны по размеру и форме, с крупной птеростигмой; голени с двумя апикальными шпорами; лапки 4-члениковые; церки отсутствуют. Некоторые авторы считают Tetrastigmoptera примитивными сеноедами и относят их к отряду Copeognatha (= Psocoptera). На основании наличия у Tetrastigmoptera более длинных мандибул, чем у сеноедов, высказано предположение, что Tetrastigmoptera представляют собой переходную форму между Panpsocoptera и Condylognatha; таксон, объединяющий Tetrastigmoptera и Condylognatha был назван Pancondylognatha Yoshizawa & Lienhard 2016. Сходство Tetrastigmoptera с сеноедами выражается в сходной форме тела и в сходном облике переднего крыла с большой пигментированной птеростигмой и характерной формой ячейки «areola postica». Однако среди Panpsocoptera эти признаки присущи только голофилетическому таксону Eupsocida, тогда как у более примитивных представителей Panpsocoptera, каковыми являются Parapsocida, птеростигма отсутствует, а форма крыльев бывает иной (рис. 9.2.5). Tetrastigmoptera не могут относится к Acercaria, поскольку их максиллы не имеют соответствующих модификаций [см. Acercaria (1)]. Они не могут относиться и к Parametabola в целом, поскольку их крыловой аппарат сохраняет бимоторность (тогда как у всех Parametabola задние крылья способны сцепляться с передними), на голенях сохраняются две вершинные шпоры (тогда как у всех Parametabola они утрачены), а лапки сохраняют 4 членика (тогда как все Parametabola имеют не более трех тарсомеров) [см. Parametabola (2), (4), (5)]. Судя по отсутствию церков, можно предположить, что Tetrastigmoptera относятся к Metabola.
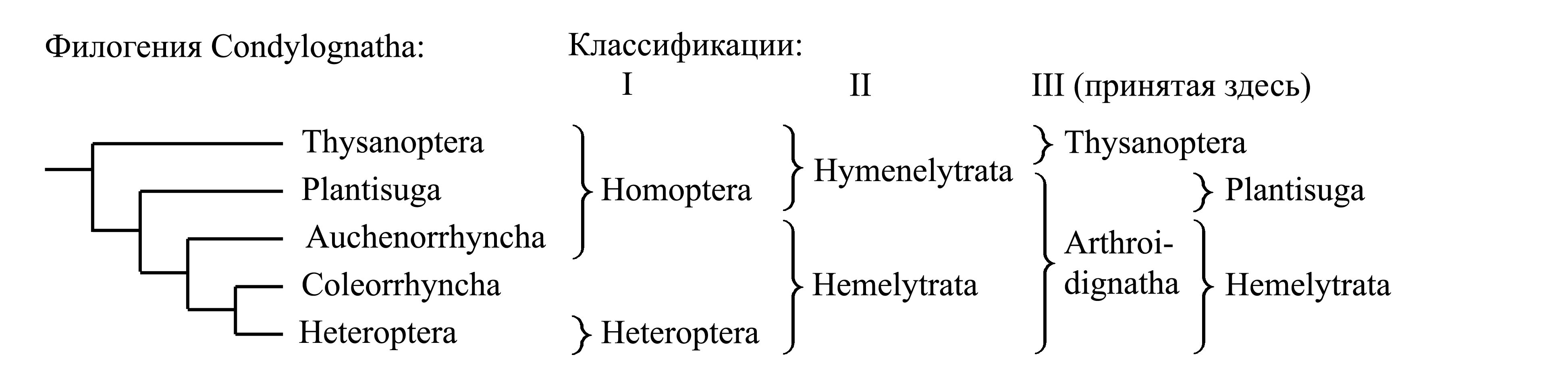
Классификации Condylognatha
I. Деление Condylognatha на Homoptera и Heteroptera. Эта классификация использовалась некоторыми авторами в первой половине XIX века. Таксон Heteroptera Latreille 1810 характеризуется аутапоморфиями и является голофилетическим (см. ниже – Heteroptera).
Таксон Homoptera Latreille 1810 {synn.circ.: Omoptera Leach 1815a = Deramphia Rafinesque 1815 = Homata Billberg 1820} является парафилетическим и характеризуется лишь отсутствием апоморфий, свойственных Heteroptera:
(1) гула отсутствует, так что ротовой аппарат [см. Condylognatha (1)] находится в задней части головы, и нижняя губа причленяется вплотную к переднегруди;
(2) переднее крыло не разделено на кориум и перепоночку.
II. Деление Condylognatha на Hymenelytrata и Hemelytrata. Эта классификация была предложена взамен предыдущей в первой половине XIX века. Здесь парафилетический таксон Homoptera расформирован таким образом, что часть его объединена с Heteroptera в голофилетический таксон Hemelytrata Fallén 1829 (см. ниже – Hemelytrata), а часть составляет собой парафилетический таксон Hymenelytrata Fallén 1829. Hymenelytrata характеризуется лишь отсутствием того способа складывания крыльев, который характерен для Hemelytrata.
III. Деление Condylognatha на Thysanoptera и Arthroidignatha. Наиболее признанным в настоящее время является деление Condylognatha на два голофилетических таксона – Thysanoptera и Arthroidignatha.
Здесь мы принимаем деление Condylognatha на два подчиненных таксона – Thysanoptera и Arthroidignatha (стр. 000).
Condylognatha, или Cimex/f1=Cicada/g3
![]() 1. Thysanoptera,
или Thrips/fg1–2
1. Thysanoptera,
или Thrips/fg1–2
![]() 2. Arthroidignatha,
или Cimex/f2=Cicada/g4
2. Arthroidignatha,
или Cimex/f2=Cicada/g4
![]() 2.1. Hemelytrata, или Cimex/f3=Cicada/g5
2.1. Hemelytrata, или Cimex/f3=Cicada/g5
![]() 2.1.1. Auchenorrhyncha, или
Cicada/f1=g6
2.1.1. Auchenorrhyncha, или
Cicada/f1=g6
![]() 2.1.1.1. Subtericornes, или
Fulgora/fg1
2.1.1.1. Subtericornes, или
Fulgora/fg1
![]() 2.1.1.2. Euhomoptera, или
Cicada/f2=g7
2.1.1.2. Euhomoptera, или
Cicada/f2=g7
![]() 2.1.2. Heteropteroidea, или
Cimex/f4=Notonecta/g1
2.1.2. Heteropteroidea, или
Cimex/f4=Notonecta/g1
![]() 2.1.2.1. Coleorrhyncha, или
Peloridium/fg1
2.1.2.1. Coleorrhyncha, или
Peloridium/fg1
![]() 2.1.2.2. Heteroptera, или
Cimex/f5=Notonecta/g2
2.1.2.2. Heteroptera, или
Cimex/f5=Notonecta/g2
![]() 2.1.2.3. †Progonocimex/fg1
2.1.2.3. †Progonocimex/fg1
![]() 2.2. Plantisuga, или Aphis/fg1
2.2. Plantisuga, или Aphis/fg1
![]() 2.2.1. Psyllaleyroda, или
Psylla/fg1
2.2.1. Psyllaleyroda, или
Psylla/fg1
![]() 2.2.1.1. Saltipedes, или Psylla/fg2
2.2.1.1. Saltipedes, или Psylla/fg2
![]() 2.2.1.2. Scytinelytra, или
Aleyrodes/fg1
2.2.1.2. Scytinelytra, или
Aleyrodes/fg1
![]() 2.2.2. Aphidococca, или
Aphis/fg2
2.2.2. Aphidococca, или
Aphis/fg2
![]() 2.2.2.1. Gynaptera, или Aphis/fg3
2.2.2.1. Gynaptera, или Aphis/fg3
![]() 2.2.2.2. Gallinsecta, или
Coccus/fg1
2.2.2.2. Gallinsecta, или
Coccus/fg1