КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
VIII-1.2.2.2.2.1. Dolichocera Bey-Bienko 1964 – длинноусые прямокрылые
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Neoptera
Neoptera![]() Rhipineoptera
Rhipineoptera![]() Tegminoptera
Tegminoptera
Saltatoria Neorchesopia![]() Dolichocera)
Dolichocera)
(рис. 8.5.1–8.5.4)
(рис. 8.5.1,
8.5.2, 8.5.3,
8.5.4)
|
Название, однозначно соответствующие по циркумскрипции: — Dolichocera Bey-Bienko 1964 (non Dolichocera Latreille 1829). По циркумскрипции также соответствует таксонам под типифицированными названиями, образованными от родовых названий Gryllus, Tettigonia и Locusta: Gryllina, Tettigonioidea, Locustina, Locustinae и др. Иерархическое название: Gryllus/f5=g3 [f:1781; g:L.1758(gen.194)] (sine Acrida; incl. Gryllacris, Tettigonia). |
|
Ранг. Обычно подотряд, реже отряд или комплекс отрядов.
Название. Этот таксон часто называют «Ensifera». В действительности название Ensifera является младшим синонимом Digastria. Таксон Digastria Graber 1875 {syn.obj.: Diplogastera Graber 1875; syn.circ: Ensifera Chopard 1920} помимо насекомых, относимых к Dolichocera, исходно включал Tridactylus/fg1 – таксон, ныне относимый к Caelifera (см. Caelifera: «Классификация»).
Аутапоморфия Dolichocera
(1) Модификация яйцеклада. Третьи створки яйцеклада сцеплены вторичными олистетерами со сцепленными между собой первыми и вторыми створками, так что весь яйцеклад двигается как единое целое; в результате этого мышцы, идущие от оснований первых и третьих створок к внутреннему гребню на боках IX тергита брюшка, служат антагонистами (мышцы «3» и «9» на рис. 8.5.2) [см. Saltatoria (7)].
Вторичный олистетер сцепляет третьи створки либо со вторыми створками, либо с первыми створками; это новообразование, возникшее у Dolichocera в дополнение к первичному олистетеру, сцепляющему первые и вторые створки первичного яйцеклада [см. Amyocerata (7)]. Вторичный олистетер устроен следующим образом (рис. 8.5.2). На третьей сворке имеется либо выступ (рис. 8.5.2A), либо продольный гребень, свободный край которого направлен медиально и загнут дорсально (так что на поперечном срезе он имеет вид крюка – рис. 8.5.2B, D, G, H). Этот выступ или гребень третьей сворки образует с первичным яйцекладом скользящее соединение, различное у Stratensifera, Mesensifera и Striduloptera (см. «Классификации» II). У Stratensifera и Mesensifera на второй створке латеральнее рахиса первичного олистетера имеется гребень, свободный край которого направлен вентрально и загнут медиально; он сцепляется с выступом или гребнем третьей сворки (рис. 8.5.2A–G). У Striduloptera на первой створке латеральнее аулакса первичного олистетера имеется гребень, свободный край которого загнут вентрально; он сцепляется с гребнем третьей сворки (рис. 8.5.2H). В обоих случаях все три створки каждой стороны (левой и правой) оказываются сцепленными между собой скользящими соединениями – первичным и вторичным олистетерами. Первичный олистетер (сцепляющий первые сворки со вторыми) сохраняется у Stratensifera, Mesensifera и Tettigensifera (хотя у Tettigensifera он становится ненужным), а у Gryllensifera он исчезает в связи с редукцией вторых створок. Сцепочный гребень на третьей сворке может тянуться вдоль нее от основания до вершины и сцеплять сворки по всей их длине; у Stratensifera-Rhaphidophora/fg1 он представляет собой короткий выступ, который сцепляет сворки только на коротком отрезке (рис. 8.5.2A). Основания левой и правой створок третьей пары жестко соединены между собой поперечным мостом. Таким образом, все шесть створок оказываются прочно соединенными вместе и образуют единый прокалывающий или режущий яйцеклад с каналом для проведения яиц. Вторичные олистетеры, как и первичные олистетеры, имеются только у имаго; у личинки недействующий яйцеклад также состоит из трех пар сомкнутых створок, но они не соединены олистетерами (рис. 8.5.2E).
Вопрос о направление эволюции вторичного олистетера вызвал разногласия, поскольку во внешней группе (т.е. у всех насекомых кроме Dolichocera) вторичного олистетера нет. По одной из гипотез вторичный олистетер исходно сцепляет третьи сворки с первыми, а у Stratensifera он вторично переместился на вторые сворки (см. «Классификации» I).
Согласно другой гипотезе вторичный олистетер исходно сцепляет третьи сворки со вторыми, а у Striduloptera он вторично переместился на первые створки (см. «Классификации» II). Этой гипотезе соответствует следующий эволюционный сценарий. Исходно третьи створки приобрели сцепление со вторыми створками; поскольку вторые створки сцеплены с первыми створками первичными олистетерами, все три пары створок оказались прочно сцепленными друг с другом (рис. 8.5.2A, B). Поскольку движение яйцеклада осуществляется антагонистическими мышцами, идущими к основаниям третьих и первых створок [см. Saltatoria (7)], а третьи и первые створки связаны между собой только опосредованно через вторые створки, все три пары створок оказываются необходимыми и не могут редуцироваться, не нарушив функционирование яйцеклада. Такое строение яйцеклада сохраняется в плезиоморфоне Stratensifera. В ходе эволюции сцепка третьих створок переместилась со вторых створок на первые. Промежуточное состояние сохраняется у Mesensifera, у которых третья сворка своим медиальным гребнем остается сцепленной со второй створкой, а своим вентральным краем входит в паз на первой сворке (рис. 8.5.2F–G). Следующим эволюционным шагом было полное перемещение вторичного олистетера, так что он стал сцеплять третьи створки непосредственно с первыми (рис. 8.5.2H). Такое состояние является исходным для Striduloptera и сохраняется у всех Striduloptera-Tettigensifera. Поскольку здесь для работы яйцеклада необходимы только сцепленные третьи и первые створки, вторые створки стали ненужными. У Tettigensifera потерявшие свое значение вторые створки сохраняются и продолжают быть сцеплены первичными олистетерами с первыми створками. У Gryllensifera вторые створки подвергаются редукции, так что в яйцекладе остаются только две развитые пары сворок – третьи и первые, сцепленные вторичными олистетерами.
Некоторые авторы усматривали сходство в строении яйцекладов Dolichocera и Notoptera, поскольку и у тех, и у других все три пары сворок неподвижны относительно друг друга, и яйцеклад двигается как единое целое. Однако у Notoptera нет вторичных олистетеров, а неподвижность вторых створок относительно третьих вызвана утратой подвижности их сочленения [ср.: Notoptera (5) и рис. 8.2.3C].
Признак Dolichocera неясной полярности
(2) Отсутствие аролия. Аролий отсутствует; иногда на его месте имеется лишь маленький выступ. По отсутствию аролия Dolichocera отличаются от Spongiphori (ср.: Caelifera: «Классификация») и от некоторых примитивных вымерших сальтаторий [ср.: Permorchesopia (2)]. Аролий имеется и отсутствует у разных насекомых, и полярность этого признака неясна [см. Pterygota (5)].
Признаки Dolichocera спорного филогенетического значения
(3) Подколенный слуховой аппарат. На голени передней ноги имеется особый слуховой аппарат – подколенный тимпанальный орган, являющийся новообразованием и не встречающийся у других насекомых (рис. 8.5.3A). Подколенный тимпанальный орган состоит из двух тимпанов (tympana), каждый из которых представляет собой участок поверхности голени с тонкой кутикулой, к которой изнутри прилегает вздутая трахея; тимпаны имеют округлую или овальную форму и расположены в проксимальной части голени на ее наружной стороне (т.е. на стороне, обращенной наружу при согнутом колене); оба тимпана располагаются на равном расстоянии от коленного сочленения, симметрично по отношению к плоскости симметрии ноги. В обычном положении передние ноги повернуты вперед, так что их тимпаны обращены вперед. Тимпаны связаны с подколенными хордотональными сенсиллами и позволяют воспринимать и распознавать звуки, издаваемые при стридуляции самцов. В связи с увеличением трахей передних ног, участвующих в образовании подколенных тимпанов, дыхальца передней пары [т.е. межсегментарные стеноторакальные дыхальца – см. Hexapoda (4b)] сильно увеличены (рис. 8.5.3A, F).
Подколенные хордотональные сенсиллы, помимо Dolichocera, найдены у многих других насекомых и, вероятно, являются исходными для насекомых; но тимпанальный орган на этом месте не встречается ни в каких таксонах кроме Dolichocera. У других Saltatoria, способных воспринимать звуки, могут быть развиты слуховые органы иного происхождения, расположенные в других частях тела; в частности, у саранчовых имеются тимпанальные органы на брюшке (ср.: Caelifera: «Классификация»: Acoustogastra).
У некоторых кузнечиков каждый тимпан защищен лопастью, образованной разрастанием края толстой кутикулы, обрамляющей тимпан сзади; в этом случае вместо овального тимпана на поверхности голени видна лишь продольная или дуговидная щель, ведущая в камеру, дно которой образует тимпан. Иногда на голени имеются не два, а лишь один тимпан.
У кузнечиков (Tettigensifera) подколенные тимпанальные органы развиты не только у имаго обоих полов, но и у личинок (рис. 8.5.3A); у сверчковых (Gryllensifera) они развиты только у имаго. У видов, утративших стридуляционный аппарат, подколенный слуховой орган либо сохраняется, либо утрачен на всех стадиях развития.
Подколенные тимпанальные органы имеются у большинства Mesensifera, Tettigensifera и Gryllensifera, но отсутствуют у всех Stratensifera (табл. VIII.2). Согласно одной из гипотез, они исходно свойственны Dolichocera, но вторично утратились у Stratensifera, так же, как и у части Mesensifera, Tettigensifera и Gryllensifera (см. «Классификации» I). Согласно другой гипотезе они исходно отсутствуют у Stratensifera и являются аутапоморфией таксона Acoustopoda, объединяющего Mesensifera, Tettigensifera и Gryllensifera (см. «Классификации» II).
(4) Тегминальный стридуляционный аппарат. У самцов на передних крыльях [которые превращены в надкрылья (тегмины) и в покое налегают друг нам друга своими клавусами – см. Saltatoria (4)] может быть развит стридуляционный (стрекотательный) аппарат следующего строения (рис. 8.5.4). Наиболее передняя из клавальных жилок (вероятно, соответствующая жилке Pcu, или A1) искривлена S-образно, а ее поперечный участок, называемый стридуляционной жилкой, утолщен и на вентральной стороне несет множество поперечных гребней, так что в профиль выглядит пиловидным (рис. 8.5.4D) и представляет собою смычок; задний край надкрылья позади стридуляционной жилки отогнут дорсально, так что на дорсальной стороне надкрылья он образует склеротизованный выступ – плектрум (plectrum). Дистальнее стридуляционной жилки продолжение Pcu ветвится и образует замкнутые петли, обрамляющие одно или несколько округлых полей, называемых зеркалами. Стридуляция осуществляется благодаря тому, что при сведении и разведении надкрылий смычок налегающего надкрылья трется о плектрум подлегающего надкрылья, а зеркало служит резонатором.
Звуки, издаваемые при стридуляции самца, воспринимаются слуховым аппаратом, имеющимся и у самцов, и у самок [см. (3)].
У кузнечиков (Tettigensifera), клавальный желоб оканчивается на заднем крае надкрылья, так что клавус со стридуляционным аппаратом занимает лишь небольшой задне-проксимальный участок надкрылья (рис. 8.5.4A). У сверчков (Gryllensifera) и профолангопсид (Prophalangopsis/fg) клавальный желоб тянется до вершины надкрылья, при этом клавус сильно расширен, так что стридуляционный аппарат занимает большую часть надкрылья (рис. 8.5.4C).
Поскольку стридуляционный аппарат расположен в проксимальной части надкрыльев, он бывает развит не только у крылатых видов, но и у многих нелетающих короткокрылых видов и у видов с укороченными надкрыльями.
Стридуляционный аппарат бывает симметричным или асимметричным. У многих кузнечиков (Tettigensifera) левое и правое надкрылья различаются по строению: смычок лучше развит на левом надкрылье, а зеркало и плектрум – на правом; в сложенном положении левое надкрылье налегает на правое, так что смычок левого надкрылья трется о плектрум правого надкрылья (рис. 8.5.4B). У профалангопсид (Prophalangopsis/fg1) и сверчковых (Gryllensifera) левое и правое надкрылья имеют одинаковое строение, и налегать может быть любое из них.
Обычно тегминальный стридуляционный аппарат развит у самцов и отсутствует у самок; у Tettigensifera-Bradyporus/fg стридуляционный аппарат имеется и у самцов, и у самок. У некоторых видов он редуцирован у самцов, что может быть связано с феминизацией строения их крыльев; иногда стридуляционный аппарат полностью утрачен.
Тегминальный стридуляционный аппарат имеется у большинства Tettigensifera и Gryllensifera, но отсутствуют у всех Stratensifera и Mesensifera (табл. VIII.2). У полнокрылых видов Stratensifera и Mesensifera надкрылья в покое складываются облегающе: в дистальной части они широко налегают друг на друга, причем каждое надкрылье плавно перегибается через среднюю линию тела и свешиваются с обратной стороны (рис. 8.5.1A) [см. Saltatoria (4)]. При таком способе складывания надкрыльев ими вообще невозможно совершать движения из стороны в сторону, необходимые для тегминальной стридуляции.
На основании ошибочной гомологизации жилок было высказано утверждение, будто стридуляционные жилки у кузнечиков и сверчков негомологичны, и их стридуляционные аппараты имеют независимое происхождение (Шаров 1968). Эта ошибка в гомологизации жилок вызвана тем, что у самцов многих кузнечиков клавусы левого и правого надкрыльев так глубоко и по-разному модифицированы и склеротизованы, что жилки на них не прослеживаются. В тех случаях, когда жилки отчетливо выражены, и у кузнечиков, и у сверчков стридуляционная жилка представляет собой непосредственное продолжение самой передней жилки клавуса – т.е. Pcu (рис. 8.5.4A, C). Поперечные гребни на стридуляционной жилке, вероятно, соответствуют экзотенидиям, имеющимся на тех или иных жилках у многих насекомых [см. Pterygota (4Ae)].
Согласно одной из гипотез, тегминальный стридуляционный аппарат исходно присущ общему предку Dolichocera, но вторично утрачен у Stratensifera и Mesensifera, подобно тому, как он утратился у части Tettigensifera и Gryllensifera (см. «Классификации» I). Согласно другой гипотезе он исходно отсутствует у Stratensifera и Mesensifera и является аутапоморфией таксона Striduloptera, объединяющего Tettigensifera и Gryllensifera (см. «Классификации» II).
Плезиоморфия Dolichocera
(5) Длинная щетинковидная антенна. Флагеллум антенны всегда щетинковидный, т.е. сужается к вершине [см. Amyocerata (1)], как правило длинный (обычно длиннее тела), с большим числом члеников (рис. 8.5.1A); членики флагеллума цилиндрические, короткие. У личинки с каждым возрастом число члеников добавляется путем деления проксимальных члеников, при этом дистальные членики могут отбрасываться [см. Amyocerata (1c)] (отбрасывание дистальной части антенны наблюдалось мною у Phaeophilacris bredoides Kaltenbach 1986).
Размеры. Длина тела 2–100 мм.
Геологический возраст. Триас – ныне (Rasnitsyn & Quicke 2002). Наиболее древними являются разнообразные триасовые виды, относимые к таксону Hagla/fg [f:1906; g:1856] (Горохов 1995); принадлежность этих насекомых к Dolichocera обосновывается наличием у самцов стридуляционного аппарата на передних крыльях.
Состав и распространение. Более 13 000 видов (Zhang 2011); всесветно.
Классификации Dolichocera
I. Деление Dolichocera на Tettigonia/g и Gryllus/g. Некоторые авторы объединяют Tettigensifera, Stratensifera и Mesensifera и противопоставляют их Gryllensifera (см. II). Эта классификация основана на предположении, что в яйцекладе вторичный олистетер исходно сцепляет третьи створки с первыми (что имеет место у Tettigensifera и Gryllensifera), а у Mesensifera и Stratensifera он изменился и стал сцеплять третьи сворки со вторыми. При этом отсутствие у всех Stratensifera и Mesensifera тегминального стридуляционного аппарата и отсутствие у всех Stratensifera подколенного тимпанального слухового аппарата объясняют их вторичной утратой (Горохов 1995).
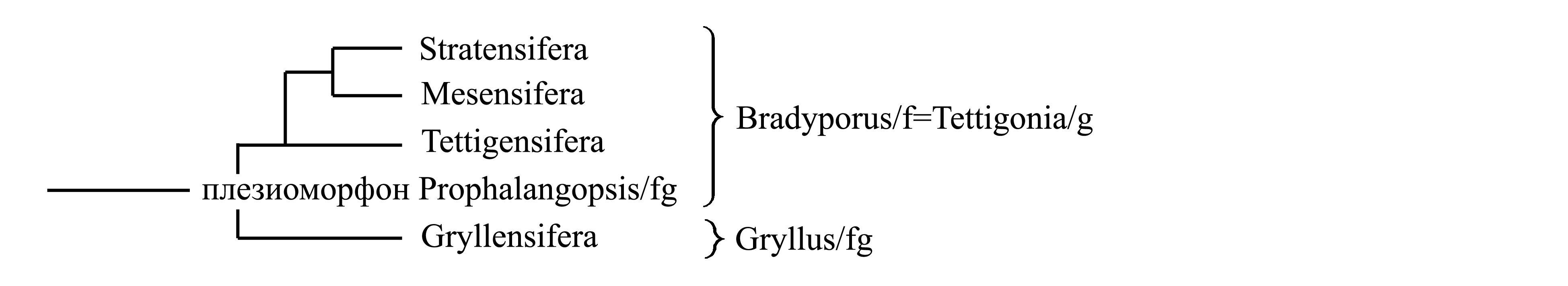
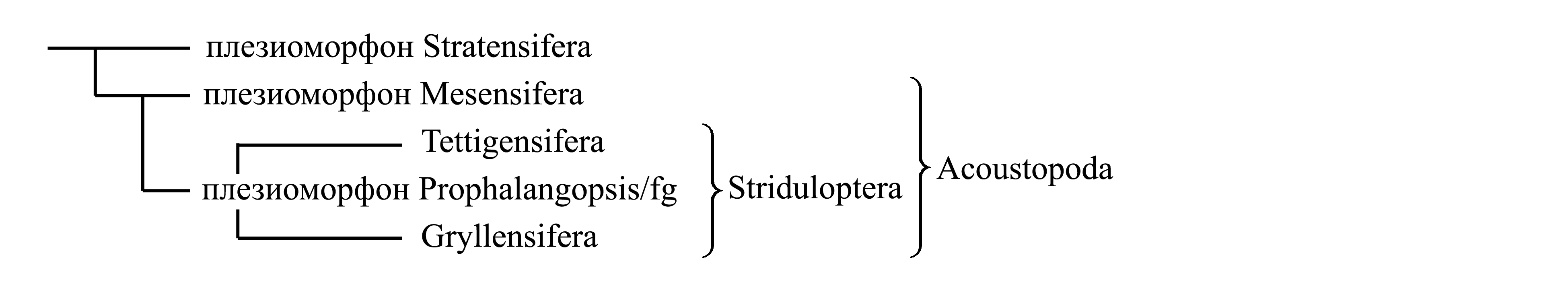
II. Выделение таксонов Acoustopoda и Striduloptera. Эта классификация основана на предположении, что подколенный слуховой аппарат и тегминальный стридуляционный аппарат не являются исходными для Dolichocera, а возникли как новообразования у таксонов Acoustopoda и Striduloptera соответственно. Согласно этому предположению, исходно Dolichocera имели лишь феморо-абдоминальный стридуляционный аппарат, унаследованный ими от общего предка Saltatoria, а звуки воспринимали внутренними слуховыми органами, не имеющими внешних тимпанов. Среди Dolichocera такое состояние сохранилось у плезиоморфона Stratensifera; то же у части Caelifera [см. Saltatoria (2b)]. У Dolichocera-Acoustopoda появились тимпанальные подколенные слуховые органы, которые исходно воспринимали звуки, издаваемые феморо-абдоминальным стридуляционным аппаратом; такое состояние сохранилось у плезиоморфона Mesensifera. Поскольку тимпанальные органы способны воспринимать более разнообразные звуки, чем слуховые органы без тимпанов, у Acoustopoda-Striduloptera появился принципиально новый способ стридуляции – тегминальный. В результате, у всех способных к стрекотанию представителей Striduloptera звуки производятся тегминальным аппаратом самца и воспринимаются подколенными тимпанальными органами. Эта гипотеза об эволюции стридуляционных и слуховых аппаратов от Stratensifera через Mesensifera к Striduloptera хорошо согласуется с гипотезой об эволюции яйцеклада от сцепки третьих сворок со вторыми к сцепке третьих створок с первыми и далее к редукции вторых сворок, согласно которой эволюция шла от плезиоморфона Stratensifera через плезиоморфон Mesensifera к Striduloptera, а внутри Striduloptera – от строения, свойственного Tettigensifera к строению, свойственному Gryllensifera [см. Dolichocera (2)].
1. Плезиоморфон Stratensifera Kluge 2016 {nom.typ. Stenopelmatus/f=Gryllacris/g [f:1838; g:1831] (incl. Rhaphidophora)}. Название «Stratensifera» («слоистомеченосные») связано с налеганием третьих створок яйцеклада на первые, что придает слоистость мечевидному яйцекладу. В яйцекладе вторичный олистетер сцепляет третьи створки не с первыми, а со вторыми сворками (то же у Mesensifera; в отличие от Striduloptera) [см. Dolichocera (1)]; при этом вентральные края третьих сворок свободные, не вставлены в пазы на первых сворках, и третьи створки налегают с боков на первые створки, закрывая их частично или полностью (рис. 8.5.1A, D, 8.5.2A–D) [в отличие от Acoustopoda (= Mesensifera + Striduloptera)]. Тимпанальные слуховые органы всегда отсутствуют (в отличие от многих Acoustopoda). Тегминальный стридуляционный аппарат всегда отсутствует (в отличие от многих Striduloptera); на переднем крыле клавальный желоб изогнут вперед, так что у полнокрылых форм надкрылья в покое складываются облегающе, в дистальной части перегибаясь на противоположную сторону (рис. 8.5.1A) (то же у Mesensifera) [см. Dolichocera (4)]. У некоторых представителей развит феморо-абдоминальный стридуляционный аппарат; в этом случае шипики на брюшке образуют вертикальные дуговидные ряды (рис. 8.5.1A) [см. Saltatoria (2b)]. Таксон Stratensifera включает около 1000 видов; в его составе принято выделять следующие подчиненные таксоны.
1.1. Rhaphidophora/fg [f:1871; g:1838] (рис. 8.5.2A–B). Все виды бескрылые; часто ведут скрытый образ жизни. Лапки сжаты с боков, без парных плантул (могут иметь непарные плантулы ); следы слияния 1-го и 2-го тарсомеров [см. Neorchesopia (1)] утрачены, так что лапки 4-члениковые. Широко распространены в тропиках и субтропиках; синантропные виды имеют космополитическое распространение.
1.2. Gryllacris/fg [f:1845; g:1831] (рис. 8.5.1, 8.5.2C–D). Многие виды способны выделять изо рта паутину, секретируемую лабиальными железами (Walker et al. 2012); обитают на деревьях и делают убежища из листьев, скрепленных паутиной. Широко распространены в тропиках и субтропиках.
1.3. Stenopelmatus/fg [fg:1838]. У бескрылых представителей средне- и заднегрудь уменьшены, так что тело выглядит суженным между переднегрудью и брюшком (у крылатых видов форма тела обычная). В Ориентальной, Афротропической, Неотропической и Неарктической областях.
Предположительно к Stratensifera относится Schizodactylus/fg [f:1845; g:1835], утративший яйцеклад. Крылья складываются веерообразно так, что образуют плотные жгуты, вершины которых скручиваются спирально вверх. В Азии и Африке.
2. Acoustopoda Kluge 2016 {nom.hier.: Gryllus/f6=g4 [f:1781; g:1758] (sine. Gryllacris; incl. Anostostoma)}. Характеризуется двумя уникальными аутапоморфиями.
(1) Появились подколенные органы слуха с тимпанами (рис. 8.5.3A) [см. Dolichocera (3)]. Они используются для восприятия звуков, издаваемых либо первичным для Saltatoria феморо-абдоминальным стридуляционным аппаратом (у Mesensifera), либо новоприобретенным тегминальным стридуляционным аппаратом (у Striduloptera). У некоторых непоющих форм органы слуха вторично утрачены.
(2) В яйцекладе вентральные края третьих створок вставлены в пазы на дорсальных краях первых сворок, так что не закрывают первые створки с боков (рис. 8.5.2F–H) [см. Dolichocera (1)]. Эта особенность проявляется только у имаго, тогда как у личинок [у которых створки протовипозитора не сцеплены олистетерами – см. Amyocerata (7)] третьи сворки сохраняют исходное положение по бокам от первых и вторых створок (рис. 8.5.2E).
Таксон Acoustopoda делится на плезиоморфон Mesensifera и голофилетический таксон Striduloptera.
2.1. Плезиоморфон Mesensifera Kluge 2016 {nom.typ: Anostostoma/fg [f:1859; g:1837]} (рис. 8.5.2E–G). Название «Mesensifera» связано со строением мечевидного яйцеклада, промежуточным между Stratensifera и Tettigensifera: в яйцекладе вторичный олистетер сцепляет третьи сворки со вторыми (как у Stratensifera), но при этом вентральные края третьих створок вставлены в пазы на первых сворках (как у Tettigensifera) (рис. 8.5.2G) [см. Dolichocera (1)]. Тимпанальные слуховые органы бывают развиты или утрачены [см. Acoustopoda (1)]. Тегминальный стридуляционный аппарат всегда отсутствует (в отличие от многих Striduloptera); на переднем крыле клавальный желоб изогнут вперед, так что у полнокрылых форм надкрылья в покое складываются облегающе, в дистальной части перегибаясь на противоположную сторону (то же у Stratensifera). У некоторых представителей развит феморо-абдоминальный стридуляционный аппарат; в этом случае шипики на брюшке расположены беспорядочно, не образуя рядов [см. Saltatoria (2b)]. Включает около 200 видов в Южной и Северной Америке, Юго-Восточной Азии, Африке, Австралии и Новой Зеландии.
2.2. Striduloptera Kluge 2016* {nom.hier.: Gryllus/f7=g5 [f:1781; g:1758] (sine Anostostoma; incl. Prophalangopsis, Tettigonia)}.
| *Бету (Bethoux 2012) предложил для этого таксона название «Grylloptera» с авторством «Haeckel 1896» и новым определением, которое он считает первым филогенетическим определением этого названия. В действительности именно Геккель ввел термины «филогения» и «филогенетическая классификация», поэтому все установленные им таксоны и их названия следует считать филогенетически определенными им самим. Название Grylloptera Haeckel 1896 является младшим циркумскриптным синонимом названия Orthoptera Olivier 1789 (см. Saltatoria: «Название»). |
Характеризуется двумя уникальными аутапоморфиями.
(1) Появилась тегминальная стридуляция самцов: по крайне мере есть стридуляционная жилка – S-образно изогнутая клавальная жилка с вентральными зубчиками (рис. 8.5.4) [см. Dolichocera (4)]; при этом звук, издаваемый самцом, воспринимается подколенными тимпанальными органами, имеющимися у обоих полов [см. Acoustopoda (1)]. У непоющих форм этот аппарат вторично утрачен.
(2) Вторичный олистетер яйцеклада изменен таким образом, что сцепляет третьи сворки не со вторыми, а непосредственно с первыми (рис. 8.5.2H). В результате этого вторые створки [которые исходно обеспечивали опосредованную сцепку третьих сворок с первыми – см. Dolichocera (2)] потеряли значение и могут подвергаться редукции; у Tettigensifera они остаются вполне развитыми, а у Gryllensifera редуцируются.
Таксон Striduloptera включает крупные голофилетические таксоны Tettigensifera и Gryllensifera; несколько видов не относятся к ним и объединяются в плезиоморфон Prophalangopsis/fg1.
2.2.1. Плезиоморфон Prophalangopsis/fg [f:1906; g:1871], или Hagla/fg [f:1906; g:1856]. Тимпанальные слуховые органы развиты [то же у Tettigensifera и Gryllensifera; в отличие от Stratensifera – см. Dolichocera (3)]. Тегминальный стридуляционный аппарат самца имеется [см. Dolichocera (4)]; в отличие от Tettigensifera, он занимает значительную часть надкрылья и сохраняет симметричность. В отличие от Gryllensifera, лапки с перетяжками на местах слияния всех первичных тарсомеров [см. Neorchesopia (1)]; при этом число подвижных тарсомеров бывает 4 или 3. Строение яйцеклада неизвестно: типовой вид Prophalangopsis obscura известен по единственному экземпляру самца из Индии; три вида Cyphoderris Uhler 1864 (обитающие в Северной Америке) и Paracyphoderris erebeus Storozhenko 1980 (на Дальнем Востоке России) короткокрылые, с вестигиальным яйцекладом. Сюда же относят большое число вымерших мезозойских видов, в том числе Hagla gracilis; в этом случае старшим типифицированным названием всего таксона оказывается не Prophalangopsis/fg, а Hagla/fg.
2.2.2. Tettigensifera Kluge 2016 {nom.typ.: Bradyporus/f=Tettigonia/g [f:1838; g:1758]} – кузнечики (рис. 8.5.2H, 8.5.3, 8.5.4A–B). Тимпанальные слуховые органы обычно развиты [см. Acoustopoda (1)]. Тегминальный стридуляционный аппарат самца обычно имеется [см. Striduloptera (1)], компактный и более или менее асимметричный: при складывании крыльев левое надкрылье налегает на правое; зеркало правого надкрылья имеет вид небольшого четко очерченного округлого прозрачного поля, окаймленного изогнутой жилкой; плектрум лучше развит на правом надкрылье, а смычок лучше развит на левом надкрылье (рис. 8.5.4A–B). При движении надкрыльев смычок, расположенный на вентральной стороне стридуляционной жилки левого (налегающего) надкрылья, трется о плектрум правого (подлегающего) надкрылья; при этом зеркало правого (подлегающего) надкрылья служит резонатором. В яйцекладе вторичный олистетер сцепляет третьи сворки не со вторыми, а с первыми (рис. 8.5.2H) [см. Striduloptera (2)]; в отличие от Gryllensifera, вторые (внутренние) створки тоже вполне развиты. Более 6000 видов, всесветно.
2.2.3. Gryllensifera Kluge 2016 – сверчковые {nom.typ.: Gryllus/f8=g6 [f:1781; g:1758] (sine Tettigonia; incl. Gryllotalpa, Myrmecophilus, Mogoplistes)} (рис. 8.5.4C–D). Лапки [исходно имеющие не более четырех подвижных члеников – см. Neorchesopia (1)] стали 3-члениковыми, при этом места слияния трех проксимальных члеников никак не выражены. Тимпанальные слуховые органы обычно развиты [см. Acoustopoda (1)]. Тело более или менее дорсовентрально уплощено, широкое [в отличие от исходного – см. Saltatoria (2)], соответственно клавусы надкрыльев, в покое лежащие на дорсальной стороне тела, так же широкие. У самца тегминальный стридуляционный аппарат обычно развит [см. Striduloptera (1)] и занимает бóльшую часть клавуса и бóльшую часть надкрылья в целом (рис. 8.5.4C); строение правого и левого надкрыльев одинаковое; у некоторых видов стридуляционный аппарат утрачен. У летающих форм задние крылья всегда длиннее надкрыльев и имеют увеличенное число складок анального веера, так что в сложенном виде выглядят как узкие гибкие жгуты, выступающие из-под надкрыльев. У яйцеклада вторые (внутренние) створки частично или полностью редуцированы (в отличие от прочих Dolichocera), а третьи сворки сцеплены с первыми вторичными олистетерами [см. Striduloptera (2)].
В составе Gryllensifera принято выделять следующие подчиненные таксоны.
2.2.3.1. Gryllus/fg (sine Gryllotalpa, Myrmecophilus, Mogoplistes) – сверчки (рис. 8.5.4C–D). Сюда относится большая часть сверчковых. Некоторые из них обитают на растениях, многие ведут напочвенный образ жизни и способны рыть норы, но не имеют такой глубоко зашедшей роющей специализации, как у медведок. Около 3000 видов, всесветно.
2.2.3.2. Gryllotalpa/fg [f:1815; g:1802] – медведки. Роющие, ведут подземный образ жизни. Передние ноги копательные, причем копательные зубцы образованы двумя вершинными шпорами голени и двумя первыми члениками 3-члениковой лапки. Яйцеклад утрачен. Несмотря на подземный образ жизни, некоторые виды сохраняют укороченные надкрылья, способные к стридуляции, и хорошо развитые задние крылья, способные к полету. Около 100 видов, всесветно.
2.2.3.3. Myrmecophilus/fg [f:1874; g:1827]. Полностью бескрылые, стридуляционные и слуховые органы утрачены. Около 100 видов, всесветно.
2.2.3.4. Mogoplistes/fg [f:1873; g:1838]. Самки бескрылые, у самца имеются только передние крылья со стридуляционным аппаратом. Тело покрыто чешуйками. Более 360 видов, всесветно.
![]() Таблица
VIII.2. Признаки таксонов в составе Dolichocera
Таблица
VIII.2. Признаки таксонов в составе Dolichocera
|
Dolichocera |
||||
| Stratensifera |
Acoustopoda |
|||
| Mesensifera |
Striduloptera |
|||
| Tettigensifera | Gryllensifera | |||
|
Подколенный тимпанальный орган |
– | ± | ± | ± |
|
Тегминальный стридуляционный аппарат |
– | – | ± | ± |
| Облегающее складывание надкрыльев (у полнокрылых форм) | + | + | – | – |
|
Феморо-абдоминальный стридуляционный аппарат |
± | ± | – | – |
| В яйцекладе 3-я створка сцеплена с ... | 2-й | 2-й | 1-й | 1-й |
| В яйцекладе 3-я створка входит в паз на 1-й сворке | – | + | + | + |
| В яйцекладе 2-я сворка утрачена | – | – | – | + |
| Число различимых члеников лапки (подвижных или синсклеротичных) | 4 | 4 | 4 | 3 |
* * *