КЛАДОЭНДЕЗИС НАСЕКОМЫХ

![]()
|
КЛАДОЭНДЕЗИС НАСЕКОМЫХ |
|||||||
|
|
|
fg | |||||
VII-1.2.1. Odonata Fabricius 1793 – стрекозы
(Somobialia![]() Eucaryota
Eucaryota![]() Metazoa
Metazoa![]() Monostomata
Monostomata![]() Haemataria
Haemataria![]() Peritoneata
Peritoneata![]() Metameria
Metameria![]()
Gnathopoda![]() Euarthropoda
Euarthropoda![]() Mandibulata
Mandibulata![]() Atelocerata
Atelocerata![]() Hexapoda
Hexapoda![]() Amyocerata
Amyocerata![]()
Pterygota![]() Metapterygota
Metapterygota![]() Odonata)
Odonata)
(рис. 7.3.1–7.3.6)
(рис. 7.3.1,
7.3.2, 7.3.3,
7.3.4, 7.3.5,
7.3.6)
|
Названия, соответствующие по циркумскрипции всем значениям Libellula/fg1–2 (т.е. и Odonatoptera, и Neodonata – см. ниже): — Odonata Fabricius 1793; — Cryptodonta Latreille 1802:T.1:364; — Odonatea Rafinesque 1815; — Paraneuroptera Shipley 1904; — Zygopteradelphia = Panzygoptera Crampton 1916a; — Odonatida Pearce 1936 — Orthomyaria Schwanwitsh 1946; — Bimotoria Schwanwitsh 1946; — Heterognatha Chen 1962 (non Heterognatha Saussure & Humbert 1872). По циркумскрипции также соответствует таксонам под родовым названием Libellula (sensu Linnaeus 1758) и производными типифицированными названиями: Libellulaedes, Libellilinae, Libellulidae, Libellulina, Libellulida, Libelluloidea, Libelluloides и др. Иерархическое название: Libellula/fg1–2 [f:1781(ord.5); g:1758(gen.207)]. |
|
Ранг. В XX веке – обычно от отряда до инфракласса; в XIX веке – обычно от семейства до подотряда; в XVIII веке – обычно род.
Название. Если рассматривать только современных представителей, то таксон Odonata выглядит четко очерченным. В этом случае все названия, перечисленные выше, однозначно соответствуют по циркумскрипции, а иерархическое название должно быть записано как Libellula/fg1. Однако описаны ископаемые палеозойские и триасовые формы, которые имеют явное родство с современными стрекозами, но не укладываются в таксон Odonata, охарактеризованный только по строению современных представителей. Эти вымершие формы вместе с современными стрекозами образуют голофилетический таксон Odonata s.l., или Odonatoptera (или Libellula/fg1), внутри которой находится подчиненный таксон Odonata s.str., или Neodonata (или Libellula/fg2). Мы не имеем возможности рассмотреть все интересующие нас признаки этих соподчиненных таксонов, так как для вымерших форм известны лишь немногие детали строения – преимущественно жилкование крыла (об особенностях ископаемых насекомых – см. главу II). Поэтому здесь характеристика стрекоз разбита на две части. В первой части – «А. Общая характеристика Odonata» рассматриваются все признаки современных стрекоз, про многие из которых неясно, относятся ли они ко всем Odonata в широком смысле, или лишь к Odonata в узком смысле слова. Во второй части – «B. Классификация Odonata с учетом вымерших групп» рассматриваются признаки, распределение которых в этих группах достоверно известно.
VII-1.2.1.A. Общая характеристика Odonata
Аутапоморфии Odonata. О некоторых их этих признаков неясно, являются ли они аутапоморфиями Neodonata или Odonatoptera (см. VII-1.2.1.B), так как для ископаемых форм они неизвестны.
(1) Метаморфоз. И личинки, и имаго специализированы как хищники, но обитают в разных средах и имеют принципиально разные приспособления для ловли добычи: личинки перешли к обитанию в воде [см. (1a)] и охотятся при помощи маски; имаго всегда имеют хорошо развитые крылья и охотятся в воздухе, схватывая добычу ногами [см. (1b)]. Помимо обычных для крылатых насекомых различий в строении птероторакса [см. (4)–(6) и] и гениталий [см. (8)–(9)], личинки и имаго стрекоз приобрели глубокие различия в строении головы, переднегруди, брюшка [см. (1b)], антенн [см. (2)], ротового аппарата [см. (3)] и каудалий см. (10)].
(1 a) Специализация личинок. Личинки [см. Pterygota (4Сa)] адаптированы к обитанию в воде (тогда как имаго всегда сохраняют наземный образ жизни); лишь у отдельных видов личинки вторично перешли к жизни во влажной почве.
Под водой личинка дышит через покровы несмотря на это у личинки сохраняются все 10 пар наружных дыхалец [см. Amyocerata (2)], из них по крайней мере стеноторакальная пара всегда сохраняет способность функционировать. В большей или меньшей степени дыхание осуществляется ректальными жабрами – отростками или складками, вдающимися в полость прямого участка (ректума) задней кишки [эктодермальной – см. Haemataria (1)]. Задняя кишка в этом месте может образовывать жаберную камеру – расширение, имеющее особенно крупные размеры у Anisoptera. Внутрь ректальных жабр могут входить ветви трахей, отходящих от вентральной и дорсальной пары продольных трахейных стволов (у Anisoptera и некоторых Zygoptera); либо ректальные жабры имеют вид небольших складок, лишенных трахей (у некоторых Zygoptera) [о модификации ректальных жабр – см. ниже, Anisopteroides (4)]. Приспособления к трахейному дыханию растворенным в воде кислородом встречаются у некоторых других насекомых, но наличие ректальных жабр является уникальной аутапоморфией Odonata.
У равнокрылых стрекоз, имеющих слабо развитую ректальную камеру, имеются, кроме того, каудальные трахейные жабры уникального строения [см. ниже, Zygoptera (1)]. У личинок таксона Euphaea/fg (относящегося к Normopteroides в составе Zygentoma), помимо ректальных и каудальных жабр, на I–VII сегментах брюшка имеется по паре придыхальцевых трахейных жабр [см. Hexapoda (9b) и Metapterygota (6)] (рис. 7.3.1A).
Высказывалось необоснованное мнение, что водный образ жизни личинок стрекоз является не апоморфией, а плезиоморфией [см. Hexapoda (9b)].
Для ловли добычи личинки используют видоизмененную нижнюю губу – маску [см. (3с)]; остальные части ротового аппарата – мандибулы и максиллы – сохраняют грызущую функцию, как и у имаго [см. (3a) и (3b)].
В остальном специализации личинок разнообразны: личинки разных таксонов стрекоз адаптированы к обитанию в разных водоемах – от стоячих вод до быстрых потоков и имеют разные приспособления для плавания [см. ниже, Zygoptera (1) и Anisopteroides (4)].
(1 b) Специализация имаго. Имаго отличаются особыми приспособлениями к охоте в воздухе (рис. 7.3.1E). Сочленение головы с переднегрудью [снабженное парой шейных склеритов – см. Pterygota (2)] приобрело особую подвижность, затылочное отверстие сильно сужено, а задняя стенка головы большая и вогнутая. Глаза увеличены, полусферические; всегда сохраняются все 3 простых глазка [см. Amyocerata (4)]. Переднегрудь уменьшена, подвижно сочленена со среднегрудью. Благодаря сильному наклону среднегруди и заднегруди [см. (4)] основания всех трех пар ног сближены между собой и приближены к голове. Голени и бедра на внутренней стороне обычно несут два ряда длинных широко расставленных шиповидных щетинок, так что образуют ловчую корзину, которая способствует схватыванию добычи на лету. Брюшко обычно очень длинное и тонкое благодаря тому, что каждый из I–IX сегментов удлинен и утончен; жесткость этих сегментов обеспечивается тем, что их тергиты имеют почти трубчатую форму, а стерниты сужены. Крылья всегда развиты, длинные, бимоторные [см. (6)].
(1 c) Онтогенез. Из яйца выходит предличинка; ее конечности и ротовые органы почти не расчленены, направлены назад, не функционируют. Сразу после выхода из яйца предличинка линяет на личинку первого возраста, у которой придатки принимают нормальное положение. У личинки первого возраста нет простых глазков (но сложные глаза всегда развиты); антенны 3-члениковые; лапки не расчленены; птероторакс [без протоптеронов – см Pterygota (4B)] с неувеличенными плейритами; иногда нет эпипрокта и церков (но всегда есть торчащие парапрокты). При дальнейшем развитии появляются простые глазки (у Zygoptera они появляются у личинки старших возрастов, у Anisoptera лишь при линьке на имаго); расчленяется флагеллум антенны [см. (2)]; расчленяется лапка [см. (7)], плейриты среднегруди и заднегруди увеличиваются и наклоняются [см. (4)]; появляются протоптероны [особой формы – см. (5)], развивается полный набор характерных личиночных каудальных придатков [см. (10)].
(2) Модификация антенн. У личинок [водных – см. (1a)] антенны имеют постоянное число члеников и разнообразную форму; у имаго [летающих – см. (1b)] антенны вестигиальные, щетинковидной формы (т.е. сужающаяся к вершине) (рис. 7.3.1E).
У личинки флагеллум исходно имеет 5 члеников (т.е. в антенне, включая скапус и педицеллус, исходно 7 члеников) [см. Amyocerata (1)] (рис. 7.3.1A); иногда число члеников флагеллума уменьшено, редко увеличено, так что бывает от 2 до 6 у разных видов. У личинки I возраста флагеллум антенны 1-члениковый (т.е. антенна 3-члениковая); в типичном случае с каждой последующей линькой число члеников флагеллума увеличивается на один благодаря делению пополам наиболее проксимального из них, пока в флагеллуме не станет 4 членика; затем последний членик делится пополам, и флагеллум становится 5-члениковым.
При превращении личинки в имаго флагеллум укорачивается путем обрыва с отходами [см. Gnathopoda (3b)]: гиподерма дистальных члеников отделяется перетяжкой от гиподермы будущего имагинального флагеллума, а затем отрывается от него и сбрасывается вместе с экзувием.
(3) Модификации ротового аппарата. Ротовой аппарат [см. Hexapoda (1a)] адаптирован к хищничеству, с особой специализацией мандибул [см. (3a)], максилл [см. (3b)] и нижней губы [см. (3c)]; голова прогнатная, сравнительно короткая. Хищная специализация свойственна и личинкам, и имаго; в то же время состав жертв и способы охоты у личинок и имаго совершенно разные: личинки обитают в воде [см. (1a)] и ловят добычу нижней губой [см. (3c)], а имаго летающие и ловят крылатых насекомых в воздухе с помощью ног [см. (1b)].
Особый характер ротового аппарата отражен в названии «Odonata» – «зубастые»: Фабрициус (Fabricius 1792–1798) разделил всех членистоногих на 11 отрядов по строению ротового аппарата, одним из которых были стрекозы.
(3 a) Мандибулы. Мандибулы [см. Mandibulata (1) и Metapterygota (2)] не имеют молы, у имаго однообразные у всех стрекоз (рис. 7.3.3B): инцизор трехзубчатый; кинетодонт (обычно принимаемый за молу) слит с корпусом, имеет 4 зубца, соединенных Z-образным гребнем (уникальная аутапоморфия). У личинок строение мандибул иное и более разнообразное: инцизор с иным числом зубцов, кинетодонт иной формы, может быть вполне развит и отделен швом от корпуса мандибулы (рис. 7.3.3A), иногда отсутствует.
(3 b) Максиллы. У личинки и имаго максиллы [см. Hexapoda (2)] имеют сходное уникальное строение (рис. 7.3.3C): максилла имеет внутреннюю (медиальную) и наружную (латеральную) доли примерно равной длины; внутренняя доля (считающаяся либо слившимися галеей и лацинией, либо просто лацинией) заостренная, с 2 продольными рядами зубцов и крепких щетинок на внутренней (медиальной) стороне; наружная доля (считающаяся либо одночлениковым щупиком, либо галеей) обычно туповершинная, иногда заостренная. Поскольку в исходной максилле насекомых мышцы и от галеи, и от щупика идут только в стипес, при наличии лишь одного из этих придатков невозможно определить по мускулатуре, является ли он галеей или щупиком; наружная доля максиллы стрекоз одночлениковая, что возможно и для галеи, и для щупика.
(3 c) Нижняя губа. У личинки и имаго нижняя губа [см. Hexapoda (3)] видоизменена уникальным образом (рис. 7.3.1A, B, E, F): помимо непарного постментума и подвижно причлененного к нему непарного образования, называемого медиальной долей (и считающегося прементумом либо результатом полного слияния прементума, глосс и параглосс), имеется пара придатков – боковых долей, подвижно причлененных к дистально-латеральным углам медиальной доли; каждая боковая доля несет подвижно причлененный изогнутый шиповидный придаток, не имеющий мускулатуры. По мнению многих авторов, латеральные доли по своему происхождению являются лабиальными щупиками, тогда как глоссы и параглоссы слиты с прементумом и не обособлены. Согласно другой версии латеральные доли являются не щупиками, а параглоссами, тогда как щупики редуцированы.
У личинки постментум и прементум (медиальная доля) сильно удлинены и сочленены подвижно, так что нижняя губа образует маску; когда маска выдвинута, ее вершина с латеральными долями выступает далеко вперед головы (рис. 7.3.1A); когда маска сложена, сочленение постментума и прементума отводится назад под грудь, а вершина маски с латеральными долями оказывается прямо под головой. Латеральные доли у личинки исходно специализированы как хватательные, и к их основаниям подходят мощные мышцы – сгибатели и разгибатели; такая маска используется для ловли добычи (у личинок Cavilabiata форма и функция маски изменены – см. ниже, Anisoptera: «Классификация»). У имаго, в отличие от личинки, постментум и прементум не удлинены, а латеральные доли не хватательные (рис. 7.3.1F). При метаморфозе длинная личиночная маска трансформируется в короткую нижнюю губу имаго путем антироста гиподермы и сопровождается частичной дегенерацией мышц (Snodgrass 1954).
(4) Модификации груди. Птероторакс [средне- и заднегрудь – см. Pterygota (4A)] имеет уникальное строение (рис. 7.3.1E, 7.3.2) [см. также ниже, Odonatoptera (1)]. Плейриты птероторакса высокие и наклонены дорсальной стороной назад (тогда как у всех прочих Pterygota они либо не наклонены, либо наклонены дорсальной стороной вперед). Благодаря этому наклону на среднегруди левый и правый эпистерны сомкнуты дорсально по средней линии, образуя срединный шов, так что отделяют короткую переднюю часть тергита, называемую здесь акротергит (7.3.2A: at2), от остальной части тергита среднегруди (7.3.2A: pt2); на заднегруди, наоборот, левый и правый эпимеры сомкнуты по средней линии вентрально и отделяют лежащий позади них участок стернита от остальной части стернита заднегруди. Тергиты средне- и заднегруди сильно уменьшены. Фрагм, характерных для птероторакса большинства Pterygota, нет, тергиты средне- и заднегруди несут по по узкой непарной аподеме, от которой назад и латерально отходит пара небольших продольных спинных мышц (у Anisoptera в заднегруди аподема и ее мышцы редуцируется); акротергит среднегруди несет еще одну непарную аподему, к которой крепятся продольные мышцы, идущие из переднегруди. Таким образом, в отличие от всех прочих Pterygota, у Odonata продольные спинные мышцы начинаются не от фрагмы на переднем крае сегмента, а от аподемы в средней его части. В отличие от большинства других Pterygota, у Odonata слабые продольные спинные мышцы с их своеобразным прикреплением и уменьшенный тергит не служат для опускания крыла, так что крылья приводятся в движение не перекрестной мускулатурой непрямого действия, а дорсовентральной мускулатурой прямого действия (см. Pterygota: «Классификации» VI); в связи с этим дорсовентральные мышцы, лежащие под увеличенными плейритами, особенно мощно развиты. Птероторакс и его мускулатура у Odonata модифицированы настолько сильно, что гомология многих мышц и склеритов у Odonata и прочих Pterygota неясна. Средне- и заднегрудь слиты неподвижно, часто шов между ними полностью утрачен (неуникальная апоморфия).
У молодой личинки плейриты средне- и заднегруди не увеличены и не сомкнуты медиально. У личинки более старших возрастов плейриты средне- и заднегруди, подобно имагинальным, высокие, наклонные. У зрелой личинки плейриты смыкаются по средней линии, разделяя тергит среднегруди, но высота плейритов меньше, чем у имаго (рис. 7.3.1A); тергиты средне- и заднегруди, подобно имагинальным, небольшие.
(5) Перевернутые протоптероны. Протоптероны [наружные крыловые зачатки личинки, отходящие от латеральных краев мезонотума и метанотума – см. Pterygota (4B)] не торчат в стороны, а направлены своими вершинами назад и лежат на спине благодаря тому, что они перевернуты вентральной стороной вверх, а костальным краем медиально; в результате этого задние протоптероны налегают на передние протоптероны (рис. 7.3.1A). При линьке кутикула на груди трескается по шву, который идет медиально лишь до оснований протоптеронов, а затем разделяется на пару швов, обходящих протоптероны по бокам [в отличие от прочих насекомых, у которых линочный шов идет медиально до заднегруди – см. Hexapoda (14)]. По видимому, перевернутое положение протоптеронов у личинок стрекоз связано с особым строением птероторакса и с наклоном плейритов [см. (4)]. Среди всех насекомых сходное положение протоптеронов имеется только у Saltatoria.
(6) Модификации крыльев. Крылья [всегда развитые – см. (1b)] имеют уникальные особенности (рис. 7.3.1E, 7.3.2A, 7.3.6), часть из которых свойственна Odonatoptera в целом, а часть – только Neodonata. Здесь дается общее описание жилкования крыльев стрекоз; о филогенетическом значении отдельных признаков см. ниже, Odonatoptera (2)–(4) и Neodonata (1)–(5).
(6 a) Причленение крыльев. В основании крыла нет мелких аксиллярных склеритов, подобных аксиллярному аппарату поденок или новокрылых [ср.: Ephemeroptera (7b), Neoptera (1)]. Вместо этого основания продольных жилок жестко укреплены на цельной радио-анальной базальной пластинке, опирающейся на плейральный столбик; кроме того, имеется еще костальная базальная пластинка, сочлененная с костальной жилкой (C) и с передним выступом плейрального столбика (рис. 7.3.2C).
(6 b) Поза покоя. В отличие от Neoptera, крылья неспособны складываться на спине: в покое они могут быть либо подняты вверх, либо распростерты в стороны. Будучи подняты вверх и сомкнуты своими дорсальными поверхностями, крылья оказываются сильно отклоненными назад соответственно наклону птероторакса [см. (4)], благодаря чему лежат вдоль брюшка (рис. 7.3.3D: самец). Такое положение крыльев стрекоз принципиально отличается от сложенных вдоль брюшка крыльев новокрылых насекомых (Neoptera) тем, что у стрекоз вентральные поверхности крыльев обращены латерально (наружу), а костальные края крыльев обращены дорсально. Такая поза покоя является исходной для стрекоз и характерна для Zygoptera и Epiophlebioptera. У разнокрылых стрекоз (Anisoptera) задние крылья резко расширены в основании и поэтому не могут быть уложены вдоль брюшка; в покое они обычно распростерты в стороны перпендикулярно телу [см. ниже, Anisoptera (3)].
(6 c) Пропорции крыльев. Крылья всегда развиты, длинные; всегда сохраняется исходная бимоторность: обе пары крыльев всегда примерно равной величины и принимают одинаково активное участие в полете, работая самостоятельно (сцепка между передним и задним крылом никогда не образуется); по форме и жилкованию передние и задние крылья могут быть совершенно одинаковыми, либо (у Anisoptera и некоторых Zygoptera) заднее крыло немного расширено и имеет несколько измененное жилкование.
(6 d) Жилкование крыльев. У самого своего основания жилки C, Sc и RA жестко соединены постоянной поперечной плечевой жилкой (vena humeralis – h).
В проксимальной части крыло либо сужено и имеет вид стебелька с уменьшенным числом продольных жилок (рис. 7.3.2A) (у Protozygoptera, Triadophlebioptera и многих Zygoptera), либо не сужено, но и в этом случае в проксимальном участке, соответствующем стебельку, число продольных жилок уменьшено (рис. 7.3.6H). В обоих случаях число продольных жилок, идущих в области стебелька и продолжающихся дистальнее этой области, сокращено до пяти; эти жилки сближены, более или менее параллельны друг другу и жестко укреплены в основании; благодаря правильному чередованию резко выпуклых и резко вогнутых жилок они образуют пучок, жесткий в продольном направлении. В этом пучке жилок позади выпуклой C, вогнутой Sc и выпуклой RA [см. Pterygota (4Ad)] сразу следует вогнутая жилка, обозначаемая нами как кубитальная (Cu), а за ней выпуклая жилка, обозначаемая нами как анальная (A) (поскольку в A трахея всегда входит не из основания крыла, а из жилки Cu через поперечную жилку Ac, существует мнение, что ход этой трахеи соответствует истинному ходу жилки A, тогда как участок A от основания крыла до впадения Ac является ложной жилкой). У стрекоз палеозойско-триасовых групп (т.е. у всех, кроме Neodonata), помимо этих пяти продольных жилок, от основания крыла между RA и Cu (которая на рис. 7.3.5 обозначена как задняя кубитальная – CuP) отходит еще одна жилка (обозначенная на рис. 7.3.5 как передняя кубитальная – CuA) – короткая выпуклая жилка, впадающая в CuP. Позади A нет жилок, отходящих от основания крыла: в стебельчатых крыльях A образует задний край стебелька, в нестебельчатых крыльях от A назад могут отходить ветви.
Там, где кончается стебелек (или соответствующий ему проксимальный участок), от выпуклой RA (идущей почти до вершины крыла) назад отходит вогнутая жилка RS; здесь же от RS отходит выпуклая медиальная жилка (M) (ее считают соответствующей передней ветви M в крыле других древнекрылых и часто обозначают как MA). Принято считать, что проксимальнее места отхождения RS и M от RA (т.е. в области стебелька) M и RS слились с RA. Жилка M у современных стрекоз неветвящаяся; среди вымерших групп она ветвится только у Meganisoptera.
Помимо этой интерпретации жилкования, существует иная (Riek & Kukalova-Peck 1984) (рис. 7.3.5C). Она основана на двух экземплярах ископаемых карбоновых насекомых, описанных как два разных вида – Eugeropteron lunatum и Geropteron arcuatum, объединяемых в таксон Geroptera (см. ниже, Палеозойско-триасовые стрекозы). Основание M считается слившимся не с RA, а с CuP; вогнутая жилка после MA – не CuP, а MP; выпуклая жилка за ней – не A, а CuA. Эта версия вызывает сомнения, поскольку основана на расположении изменчивых косых жилок у единичных экземпляров.
Жилка RS разветвляется, образуя триаду с двумя вогнутыми ветвями – RSa и RSp (часто обозначаемыми как R2+3 и R4+5) и выпуклой интеркалярной жилкой между ними – iRS (часто обозначаемой как IR3); передняя ветвь этой триады разветвляется, образуя еще одну триаду с двумя вогнутыми ветвями – RSa1 и RSa2 (часто обозначаемыми как R2 и R3) и выпуклой интеркалярной iRSa (часто обозначаемой как IR2) [об используемых здесь обозначениях ветвей и интеркалярных – см. Pterygota (4Ad)]. Такое же ветвление RS у Ephemeroptera, что можно расценить как синапоморфию или симплезиоморфию (см. Ephemeroptera: «Систематическое положение» II).
У всех Neodonata и некоторых других стрекоз Sc идет вдоль переднего края крыла и, далеко не доходя до вершины крыла, впадает в C; место слияния Sc и C образует характерный элемент крыла стрекоз – узелок, или нодус (nodus) (N): передний край крыла в этом месте имеет надлом, а Sc соединена с RA постоянной поперечной жилкой, называемой узелковой (vena nodalis) (n); RA соединена с RSa постоянной поперечной жилкой, кажущейся продолжением узелковой жилки и называемой подузелковой (vena subnodalis) (sn).
Между плечевой жилкой и узелком обычно имеются еще две постоянные поперечные жилки, называемые первичные предузелковые жилки (venae antenodales) (an1 и an2); они соединяют C, Sc и RA. Иногда [например, у Libellula/fg (sine Cordulia), Calopteryx/fg и Euphaea/fg] первичные предузелковые бывают неразличимы среди множества вторичных предузелковых жилок (рис. 7.3.1E).
У большинства стрекоз в дистальной части крыла между RA и C имеется склеротизованный участок – птеростигма (pterostigma) (pt).
Жилки RS и M в месте их отхождения от RA имеют короткий общий стволик, обычно направленный поперек крыла и напоминающий поперечную жилку. Почти у всех современных стрекоз этот развилок связан постоянной поперечной жилкой с Cu, так что эта поперечная жилка вместе с общим стволиком RS+M (а иногда также с коротким отрезком M) образуют еще один характерный элемент крыла стрекоз – дужку (arculus) (Arc). Дистальнее дужки M соединена с Cu еще одной постоянной поперечной жилкой; между ней, дужкой, M и Cu заключена характерная для стрекоз постоянная ячейка, называемая дискоидальной (dc). У равнокрылых стрекоз (Zygoptera) дискоидальная ячейка также называется четырехугольником (quadrangulum) (q) (рис. 7.3.2A), а у разнокрылых стрекоз она разделена на треугольник (triangulum) (t) и надтреугольник (supratriangulum) (st) [см. ниже, Anisoptera (2) и рис. 7.3.6G–H].
Между названными продольными жилками может находиться большее или меньшее число дополнительных ветвей, интеркалярных и поперечных жилок, либо архедиктион – сеть жилок, образующих многоугольные ячейки (рис. 7.3.6H).
(7) Трехчлениковая лапка. Лапка [исходно пятичлениковая – см. Amyocerata (5)] по крайней мере у имаго современных стрекоз всегда 3-члениковая, первый членик самый короткий, третий – самый длинный (рис. 7.3.1E). У личинки I возраста лапка не расчленена; при последующих личиночных линьках сначала обособляется первый членик, затем разделяются второй и третий (иногда на передних и средних ногах второй и третий членики не разделяются, и лапки зрелой личинки остаются 2-члениковыми). Трехчлениковая лапка – неуникальная апоморфия (см. Указатель признаков [2.50]: табл. E). Вероятно, в данном случае это аутапоморфия Neodonata, так как у некоторых вымерших стрекоз, относящихся к Protozygoptera и Meganisoptera, известны 4-члениковые лапки.
Лапка личинки и имаго всегда заканчивается двумя коготками претарсуса, без какого-либо срединного придатка [см. Pterygota (5)].
(8) Вторичный копулятивный аппарат самца. У имаго самца имеется вторичный копулятивный аппарат, не имеющий гомологов у других насекомых (рис. 7.3.1E–G, 7.3.4C): на II стерните брюшка имеется пара передних зацепок (hamuli anteriori), служащих для фиксации створок яйцеклада самки, и пара задних зацепок (hamuli posteriori); эти зацепки сидят по краям глубокой медиальной впадины; дно этой впадины образовано сложной подвижной системой склеритов и мембран, от него отходит подвижно причлененный непарный отросток – лигула (ligula); на III стерните брюшка имеется выступающая вперед семенная капсула (vesica spermalis), способная вкладываться во впадину на II стерните. В семенной капсуле хранится сперма, помещенная сюда из полового отверстия [находящегося на IX стерните – см. (9)]. Для помещения спермы в семенную капсулу самец подгибает брюшко, касаясь девятым уромером третьего. В разных таксонах стрекоз задние зацепки, лигула и семенная капсула развиты по-разному и по-разному используются при копуляции [см. ниже, Zygoptera (2), Anisoptera (1) и Epiophlebia (1)]. В процессе копуляции самец удерживает самку каудальными хватательными придатками [см. (10)] за переднеспинку (у Zygoptera) или за голову (у Anisopteroides), а самка совмещает свои гениталии с вторичными гениталиями самца (рис. 7.3.3D, 7.3.4A–D). У личинки самца наружные зачатки вторичного копулятивного аппарата отсутствуют.
(9) Редукция первичного копулятивного аппарата самца. Первичный половой аппарат самца [исходно находящийся между IX и X сегментами брюшка – см. Amyocerata (6)] смещен вперед на IX стернит брюшка. Он состоит из пары вестигиальных унистилигеров без стерностилей, между ними располагается вестигиальный непарный пенис с половым отверстием (рис. 7.3.3E); функционирующего копулятивного аппарата на IX сегменте нет [спаривание осуществляется вторичным половым аппаратом на II–III стернитах – см. (8)]. У зрелых личинок самцов имеются наружные зачатки унистилигеров (рис. 7.3.1A).
(10) Модификации X уромера. У личинки и имаго последний, десятый сегмент брюшка модифицирован следующим образом. Латеральные части тергита распространились на вентральную сторону сегмента и слились между собой, так что тергит имеет вид цельного кольца, а парапрокты утратили соединение с IX уромером и отделены от него тергитом X уромера [то же у некоторых других насекомых – см. Amyocerata (8b)]; при этом парапрокты могут иметь вид подвижно причлененных выростов, так что сзади к кольцу, образованному тергитом, причленены 5 одночлениковых каудальных придатков – это пара парапроктов [см. (10a)], непарный эпипрокт [см. (10b)] и пара церков [см. (10c)] (рис. 7.3.1C).
(10 a) Парапрокты. У личинок всех возрастов парапрокты [отделенные от IX уромера разросшимся тергитом X уромера – см. (10)] имеют вид выступающих подвижных каудальных придатков: у современных стрекоз они либо несут парные каудальные жабры [см. ниже, Zygoptera (1) и рис. 7.3.1A], либо образуют парные створки анальной пирамиды [см. ниже, Anisopteroides (4) и рис. 7.3.4F] (тогда как у других насекомых это невыступающие склериты стенки тела). У имаго парапрокты обычно не выступающие, но у самцов многих Zygoptera образуют пару подвижных вентральных придатков, которые используются при фиксации самки (рис. 7.3.3E). К основаниям парапроктов, помимо продольных мышц, идущих от тергита десятого уромера и соответствующих исходным дорсовентральным мышцам, крепятся продольные мышцы, идущие от стернита девятого уромера (рис. 7.3.1D), что указывает на происхождение этой пары каудальных придатков [см. Amyocerata (8b)].
(10 b) Эпипрокт. У личинок эпипрокт также имеет вид выступающего каудального придатка: у современных стрекоз он либо несет непарную каудальную жабру [см. ниже, Zygoptera (1) и рис. 7.3.1A], либо образует непарную створку анальной пирамиды [см. ниже, Anisopteroides (4) и рис. 7.3.4F]. Эпипрокт бывает развит у личинок всех возрастов или не развит лишь у самых молодых личинок. У имаго эпипрокт не выступающий, но у самцов Anisopteroides несет придаток, используемый при фиксации самки [см. ниже, Anisopteroides (1) и рис. 7.3.4E].
(10 c) Церки. У личинок и имаго церки [только парные – см. Metapterygota (4)] всегда одночлениковые. У личинки церки имеют меньшие размеры, чем парапрокты и эпипрокт (рис. 7.3.1C, 7.3.4F), у самых молодых личинок могут отсутствовать. У имаго церки всегда развиты у обоих полов; у самцов всегда используются для фиксации самки при спаривании [см. ниже, Zygoptera (5) и рис. 7.3.3E; Anisopteroides (1) и рис. 7.3.4E].
Некоторые авторы (Tillyard 1917 и др.) предлагали иную гомологизацию названных пяти каудальных придатков: «церками» считали наиболее вентральные парные придатки (которые в действительности являются парапроктами), а более дорсальные парные придатки (являющиеся церками) считали вторичными копулятивными органами – «церкоидами». Такая версия основана на ошибочном «биогенетическом законе», согласно которому в ходе онтогенеза более древние по происхождению придатки (в данном случае церки) должны появляться раньше и функционировать на более ранних стадиях развития, чем эволюционно более молодые придатки (см. раздел I.4.2.1); у личинок стрекоз, в отличие от имаго, парапрокты и эпипрокт крупнее церков и появляются раньше их. На истинную гомологию каудальных придатков стрекоз указывает то, что к основаниям церков подходят только мышцы, начинающиеся от десятого тергита брюшка, а к основаниям парапроктов подходят также мышцы, начинающиеся от девятого стернита брюшка (рис. 7.3.1D) [см. Amyocerata (8)].
Плезиоморфия
(11) Первичный яйцеклад. Яйцеклад обычно имеется (рис. 7.3.3F), состоит из двух пар кинетапофизов, сцепленных олистетерами и соединенных в основании посредством пары гонангулов (вальвиферов) [см. Amyocerata (7)]; IX уромер несет пару удлиненных унистилигеров (вальв), каждый с неспециализированным стерностилем на вершине [мышца, идущая к стерностилю утрачена – см. Metapterygota (5)]. У зрелых личинок самок имеются наружные зачатки VIII и IX кинетапофизов и IX унистилигеров с неотчлененными зачатками стерностилей. В некоторых независимых друг от друга таксонах Anisoptera яйцеклад подвергся редукции.
Геологический возраст. См. отдельно Odonatoptera и Neodonata. Если принимать таксон Odonata в широком смысле (т.е. как Odonatoptera), то он имеет карбоновый возраст, древнейший среди крылатых насекомых; если принимать его в узком смысле (как Neodonata), то он имеет лишь мезозойский возраст.
Систематическое положение Odonata. Кроме принятого здесь отнесения Odonata непосредственно к Metapterygota, предлагались следующие классификации.
I. Отнесение Odonata непосредственно к Pterygota (см. Pterygota: «Классификации» II, VI и VIII).
II. Объединение Odonata, Ephemeroptera и Protorrhynchota в таксон Palaeoptera (см. Pterygota: «Классификации» IV).
III. Объединение Odonata и Ephemeroptera в таксон Subulicornes (см. Ephemeroptera: «Систематическое положение» II).
IV. Объединение Odonata, Ephemeroptera и Plecoptera в таксон Arkiptera (см. Ephemeroptera: «Систематическое положение» III).
V. Объединение Odonata и Protorrhynchota в таксон Plagioptera (см. Pterygota: «Классификации» VII).
VII-1.2.1.B. Классификация Odonata с учетом вымерших групп
Филогения Odonata представляется следующим образом:
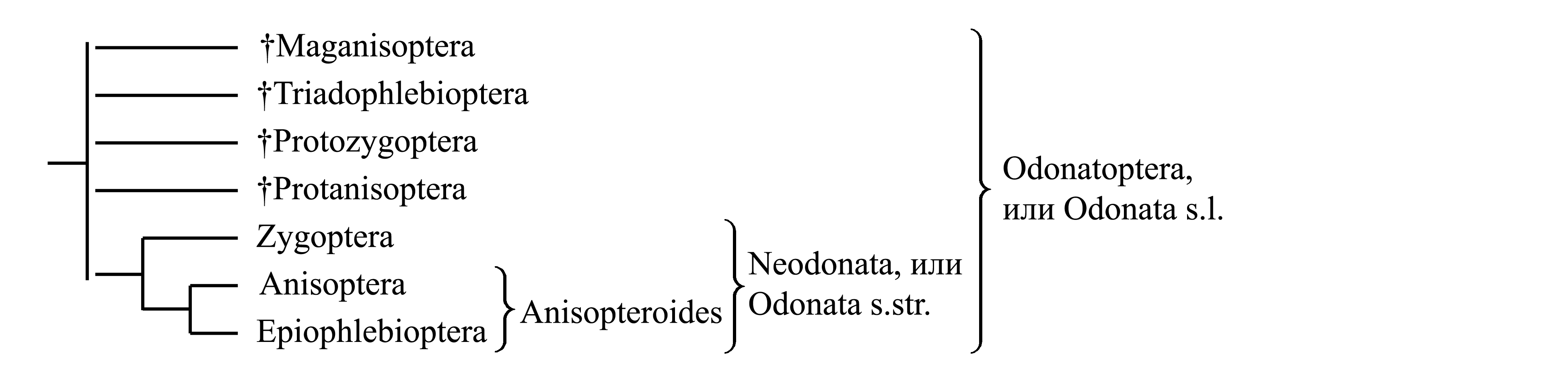
Ниже рассматривается следующая классификация стрекоз:
1. Odonatoptera, или Odonata s.l. (или Libellula/fg1)
1.1. † Палеозойско-триасовые стрекозы (стр. 0000)
1.2. Neodonata, или Odonata s.str. (или Libellula/fg2) (стр. 0000)
1.2.1. Zygoptera (или Calepteryx/f1=Lestes/g1) (стр. 0000)
1.2.2. Anisopteroides (или Libellula/fg3) (стр. 0000)
1.2.2.1. Anisoptera (или Libellula/fg4) (стр. 0000)
1.2.2.2. Epiophlebioptera (или Epiophlebia/fg1) (стр. 0000)
1.2.2.3. Вымершие группы, относящиеся к Anisopteroides (стр. 0000)
![]() 1 Odonatoptera, или Odonata
s.l. (или Libellula/fg1)
1 Odonatoptera, или Odonata
s.l. (или Libellula/fg1)
![]() 1.1 †Палеозйско-триасовые стрекозы
1.1 †Палеозйско-триасовые стрекозы
![]() 1.2 Neodonata, или Odonata s.str. (или Libellula/fg2)
1.2 Neodonata, или Odonata s.str. (или Libellula/fg2)
![]() 1.2.1 Zygoptera (или
Calepteryx/f1=Lestes/g1) – равнокрылые стрекозы
1.2.1 Zygoptera (или
Calepteryx/f1=Lestes/g1) – равнокрылые стрекозы
![]() 1.2.2 Anisopteroides (или Libellula/fg3)
1.2.2 Anisopteroides (или Libellula/fg3)
![]() 1.2.2.1 Anisoptera (или Libellula/fg4) – разнокрылые стрекозы
1.2.2.1 Anisoptera (или Libellula/fg4) – разнокрылые стрекозы
![]() 1.2.2.2 Epiophlebioptera (или
Epiophlebia/fg1)
1.2.2.2 Epiophlebioptera (или
Epiophlebia/fg1)
![]() 1.2.2.3 Вымершие группы, относящиеся к Anisopteroides
1.2.2.3 Вымершие группы, относящиеся к Anisopteroides